◄ Carnets Geol. 26 (9) ►
![]()
Outline
[1. Introduction]
[2. Methodology]
[3. Geology of Upper Eocene to lower Miocene rocks of Jamaica and their biozonation]
[4. Miogypsinid palaeontology]
[5. Evolution of the miogypsinids]
[6. Conclusions]
[Bibliographic references] [Plates] and ...
[Appendix]
Department
of Geography and Geology, The University of the West Indies, Mona, Kingston (Jamaica)
;
Published online in final form (pdf) on May 25, 2026
DOI 10.2110/carnets.2026.2609
![]()
[Editor: Alberto Collareta;
technical editor: Bruno
R.C. Granier]
![]()
Fourteen miogypsinid populations (ground-down free specimens, thin sections, and polished slabs) from Jamaica are analysed using univariate and bivariate statistics. The populations consist of one sample of free specimens from which orientated equatorial sections were prepared and thirteen populations with random equatorial sections on polished blocks. The populations are sorted into chronospecies based on mean X (Xm) values with four chronospecies (Miogypsinoides complanatus, Miogypsina thalmanni, Mio. 'basraensis', and Mio. tani) identified. Three samples collected from a single traverse show a succession of three successive chornospecies (Ms. complanatus, Mio. thalmanni, and Mio. 'basraensis'). Ms. complanatus is calibrated to around the Rupelian/Chattian boundary (based on data from Antigua), whereas advanced forms of Mio. tani are calibrated with the latest Chattian (based on planktic foraminifers, calcareous nannofossils, and Sr isotopes). The zonation for the Chattian using Larger Benthic Foraminifers is revised and five zones based on miogypsininids are recognized. This zonation enables the unconformity between the Moneague Formation (Rupelian to mid Chattian) and Newport Formation (latest Chattian) in Jamaica to be quantified. Comparison between the Americas and the Neotethys/Indo-Pacific indicates that miogypsinid evolution in the Chattian was more rapid in the Americas and that by the base of the Miocene, American miogypsinids were two chronospecies more advanced compared with their allies in the Neotethys/Indo-Pacific. This demonstrates that high resolution scanning of polished slabs represents a valuable tool for biostratigraphy of Larger Benthic Foraminifers.
• Miogypsina;
• Miogypsinoides;
• White Limestone;
• biostratigraphy;
• phylogeny;
• paleogeography
Mitchell S. (2026).- A high-resolution biostratigraphy for the Upper Oligocene (Chattian) of Jamaica using miogypsinid foraminifers, and its stratigraphic and phylogenetic significance.- Carnets Geol., Madrid, vol. 26, no. 9, p. 177-209. DOI: 10.2110/carnets.2026.2609
Une biostratigraphie � haute r�solution de l'Oligoc�ne sup�rieur (Chattien) de Jama�que fond�e sur les foraminif�res miogypsinid�s, et sa signification stratigraphique et phylog�n�tique.- Quatorze associations de miogypsinid�s (sp�cimens libres meul�s, lames minces et plaques polies) provenant de Jama�que sont analys�es � l'aide de statistiques univari�es et bivari�es. Chaque association correspond � un pr�l�vement de sp�cimens libres � partir duquel des sections �quatoriales orient�es ont �t� pr�par�es, ainsi que treize associations comportant des sections �quatoriales al�atoires sur des blocs polis. Les associations sont class�es en chronoesp�ces sur la base des valeurs moyennes de X (Xm), quatre chronoesp�ces �tant identifi�es : Miogypsinoides complanatus, Miogypsina thalmanni, Mio. 'basraensis' et Mio. tani. Trois �chantillons pr�lev�s le long d'une m�me coupe stratigraphique montrent une succession de trois chronoesp�ces successives (Ms. complanatus, Mio. thalmanni et Mio. 'basraensis'). Ms. complanatus est dat�e aux environs de la limite Rup�lien/Chattien (sur la base de donn�es provenant d'Antigua), tandis que les formes �volu�es de Mio. tani sont attribu�es au Chattien terminal (d'apr�s les foraminif�res planctoniques, les nannofossiles calcaires et les isotopes du Sr). La zonation du Chattien fond�e sur les Grands Foraminif�res Benthiques est r�vis�e, et cinq zones bas�es sur les miogypsinid�s sont reconnues. Cette zonation permet de quantifier la discordance entre la Formation de Moneague (Rup�lien � Chattien moyen) et la Formation de Newport (Chattien terminal) en Jama�que. Une comparaison entre les Am�riques et la r�gion N�ot�thys/Indo-Pacifique indique que l'�volution des miogypsinid�s durant le Chattien a �t� plus rapide dans les Am�riques et que, � la base du Mioc�ne, les miogypsinid�s am�ricains �taient avanc�s de deux chronoesp�ces par rapport � leurs homologues de la r�gion N�ot�thys/Indo-Pacifique. Cela montre que la num�risation haute r�solution de plaques polies constitue un outil pr�cieux pour la biostratigraphie des Grands Foraminif�res Benthiques.
• Miogypsina ;
• Miogypsinoides ;
•
White Limestone ;
• biostratigraphie ;
• phylog�nie ;
• pal�og�ographie
The biostratigraphic subdivision of Oligocene and lower Miocene shallow-water, platform limestones in the Americas is difficult. While planktic foraminifers and calcareous nannoplankton represent the baseline for standardized Cenozoic biostratigraphy, they are difficult to study in indurated limestones and may be absent due to the character of the depositional environments. Larger benthic foraminifers (LBF) offer significant scope for biostratigraphy using objective criteria (Mitchell et al., 2022, 2024), but are difficult to extract and identify in indurated limestones lacking orientated thin sections. This paper uses a method (polished slabs) to investigate LBF (specifically miogypsinids) in indurated rocks, calibrates LBF biostratigraphy against chronostratigraphy using key samples, and uses this to explore geological problems in Jamaica and review the phylogenetic evolution of the miogypsinids across the Americas, Neotethys, and Indo-Pacific provinces.
An understanding of the geology of the Oligocene and lower Miocene rocks in Jamaica has been achieved through geological mapping and the investigation of samples in the field and in the laboratory. Field mapping was undertaken along road and path transects with rock lithologies and bedding recorded in a notebook against GPS coordinates. A hand held GPS unit was used to record GPS positions. Samples of limestones and chalks were broken off with a 4 lb (1.8 kg) lump hammer, wetted with water, and examined in the field with a 10x hand lens. The lithology was recorded using an extended Dunham classifications (Dunham, 1962; Embry & Klovan, 1971; Lokier & Al Junaibi, 2016) and bioclasts were identified. Larger benthic foraminifers (LBF) were identified to genus and species level (where possible) in the field. Samples (rock specimens and bulk samples of unlithifield sediment) were collected for subsequent laboratory examination.
GPS data were used to construct geological maps in the drawing programme CanvasX19. Stratigraphic boundaries and faults were determined through geological mapping of lithologies and breaks in biostratigraphy. Some faults were visible in remote sensing imagery (aerial photographs and satellite imagery). Other faults (some of them with large offsets) had no representation on such imagery as they had not been active recently or fault planes had not been enhanced through geomorphological modification (limestone dissolution, etc.). Furthermore, some features on remote imagery do not represent faults and are related to other geomorphological features, such as, uplifted marine terraces (modified by karst processes) formed during the progressive uplift of Jamaica. Simplified geological maps of Jamaica and the area around Ulster Spring (parish of Trelawny) are presented here
Unlithified sediment samples were disaggregated by drying and soaking in water. If this was not successful, samples were put through several cycles of freezing and thawing in water to break them down. The broken down samples were sieved through 1 mm and 500 µm sieves, and LBF (and other bioclasts) were picked from the sieved residues. LBF were sorted into morphotypes and orientated equatorial and axial sections of individual LBF were prepared by careful grinding on a glass plate.
For lithified rock samples, standard 32 µm-thick thin sections were prepared as well as numerous polished rock slabs. Polished rock slabs were serially ground using progressively finer silicon carbide grits with a final polish using 1200 grade silicon carbide powder. Polished slabs were scanned at 6400 dpi and the scans searched for appropriate sections (equatorial and axial) of LBF. Because of the large surface area of the rock slabs, numerous approximately equatorial sections of miogypsinids (and other foraminifers) were located. Photomicrographs of LBF on the thin sections and slabs were then imaged using microscope cameras.
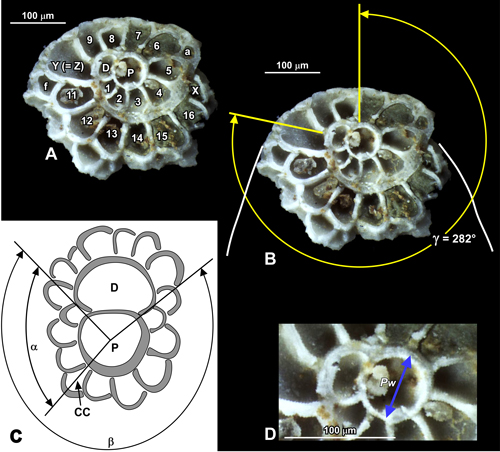
Several
characters have traditionally been measured on miogypsinids following Drooger's
(1952) statistical analysis of American populations (Fig. 1 ![]() ). These characters
were analysed by statistical methods. Since the measurements made include
categorical data (counts of numbers of chambers), with variable samples sizes,
which may not be normally distributed, nonparametric univariate statistical
tests are appropriate. Kruskal-Wallis rank sum tests for multiple
independent samples were undertaken with post-hoc Dunn pairwise tests
with p-values adjusted using the Benjamini-Hochberg
FDR method (Benjamini & Hochberg,
1995). Univariate data are
illustrated using dot plots and bar graphs. Bivariate analyses were investigated
through graphical methods with fields for samples outlined by rounded polygons.
The following sections describe the different characters that have been used for
miogypsinids (Drooger, 1952).
). These characters
were analysed by statistical methods. Since the measurements made include
categorical data (counts of numbers of chambers), with variable samples sizes,
which may not be normally distributed, nonparametric univariate statistical
tests are appropriate. Kruskal-Wallis rank sum tests for multiple
independent samples were undertaken with post-hoc Dunn pairwise tests
with p-values adjusted using the Benjamini-Hochberg
FDR method (Benjamini & Hochberg,
1995). Univariate data are
illustrated using dot plots and bar graphs. Bivariate analyses were investigated
through graphical methods with fields for samples outlined by rounded polygons.
The following sections describe the different characters that have been used for
miogypsinids (Drooger, 1952).
|
|
Figure 1: Characters measured on miogypsinids following Drooger (1952). The
illustrated specimen in A, B, and C is Miogypsinoides
cf. formosensis (image from https://foraminifera.eu
used with permission), and shows a small additional equatorial chamber (a) that
is not part of the fan of equatorial chambers (cf. chambers developed in Neorotalia).
A. chambers numbered in the primary spire with number of chambers (excluding the
protoconch [P] and the deuterioconch [D]), here X = 17. The largest chamber Z =
10 and the first chamber with two stolons that gives rise to the fan Y = 11
(these
are more subjective than determining X). B. Measurement of the angle γ: the fan of equatorial chambers is broken off, but indicated by the
white lines; angle γ
is the amount in degrees that the P-D axis needs to be rotated to 'unwind the
coil' such that D faces the apex (in this case γ
= 282�). C. Measurement of angles α and β to determine V, where V = 200α/β.
D. Measurement of the width of the protoconch Pw (including half the thickness of
the wall). |
Character
X was introduced by Drooger (1952) and is the number of chambers
(excluding
the protoconch (P) and deuterioconch (D) - that is the embryo) in the neorotalid
coil around the embryo of a miogypsinid (Fig. 1.A ![]() ). Calculated mean values (Xm)
are considered the most import characters in distinguishing populations of Miogypsinoides and early Miogypsina,
although the genera themselves are distinguished by the lack (in the former) and
presence (in the latter) of lateral chamberlets (e.g., Drooger, 1952,
1963,
1993).
However, the value of Xm
for distinguishing chronospecies of Miogypsina
is diminished once two spires of periembryonic chambers (from one or two
Principal Auxillary Chambers - PACs) encircling the protoconch and meeting in a
closing chamber are developed (Fig. 1.C
). Calculated mean values (Xm)
are considered the most import characters in distinguishing populations of Miogypsinoides and early Miogypsina,
although the genera themselves are distinguished by the lack (in the former) and
presence (in the latter) of lateral chamberlets (e.g., Drooger, 1952,
1963,
1993).
However, the value of Xm
for distinguishing chronospecies of Miogypsina
is diminished once two spires of periembryonic chambers (from one or two
Principal Auxillary Chambers - PACs) encircling the protoconch and meeting in a
closing chamber are developed (Fig. 1.C ![]() ). The first limitation on character X is
that it can only be measured if equatorial sections are available. When
matrix-free specimens are available, suitable equatorial sections can be
prepared (by grinding) so that this character can be measured. If only hard rock
samples are available, then accidental sections in thin sections or on polished
rock slabs (as used here) must be used. In the latter case, limitations are
introduced, because an informed decision needs to be made for each random
section on whether the section shows the complete neorotalid spire. Small errors
here are not going to significantly affect the mean if a 'moderate'-sized
population (8 or more specimens) is available. Counts on just one or two
specimens are of limited biostratigraphic value, since they could belong to one
of several successive chronospecies.
). The first limitation on character X is
that it can only be measured if equatorial sections are available. When
matrix-free specimens are available, suitable equatorial sections can be
prepared (by grinding) so that this character can be measured. If only hard rock
samples are available, then accidental sections in thin sections or on polished
rock slabs (as used here) must be used. In the latter case, limitations are
introduced, because an informed decision needs to be made for each random
section on whether the section shows the complete neorotalid spire. Small errors
here are not going to significantly affect the mean if a 'moderate'-sized
population (8 or more specimens) is available. Counts on just one or two
specimens are of limited biostratigraphic value, since they could belong to one
of several successive chronospecies.
Some authors have used a typological approach for the study of miogypsinids. For instance, Boudagher-Fadel and Price (2010, p. 574) stated that for Mio. gunteri, X = 10 to 12, and for Mio. tani, X = 8. Yet, Drooger (1952, 1993) clearly stated that using mean values for X (that is Xm), Mio. gunteri was defined as having Xm between 10.5 and 9 (with a range of X-values from 14 to 8 - taken from Drooger's, 1952, Table 1), whereas Mio. tani was defined as having a Xm of less that 9 (and a range of X-values from 10 to 6 - taken from Drooger's, 1952, Table 1). Thus, individual specimens cannot be assigned to a particular chronospecies, only populations with a sufficient number of specimens. Drooger's (1952) figured specimens of Mio. tani showed X = 7 to 9 chambers (his plate 2, five drawings) and one photomicrograph (on his Plate 3) showing 7 chambers. It is, therefore, difficult to understand Boudagher-Fadel and Price's (2010) statement that for Mio. tani X = 8. Consequently, their typological identification of specimens assigned to either Mio. gunteri or Mio. tani (or for that matter, other miogyspsinid species) is suspect and their biostratigraphic conclusions are open to question. Equally, Baumgartner et al. (2008, p. 40) stated that "we can clearly identify the morphological characters of the two species Mio. gunteri and Mio. tani", and that "the X-value of Miogypsina tani ranges from 6 to 9 and that of Miogypsina gunteri from 9 to 12.5". Yet these 'ranges' should be mean values of X (Xm) and, as such, individual specimens should not be assigned to chronospecies. So identifications of miogypsinid taxa in the Americas using X-values (rather than Xm values) post-Drooger (1952) have been based on species names applied to individual specimens and, consequently, have limited biostratigraphic value.
Character
Y (Fig. 1.A ![]() ) represents the number of chambers in the neorotalid spire
(excluding
P and D) up to, but NOT including the first chamber that gives rise to the fan
of equatorial chambers (Drooger, 1952; Raju,
1974). Character Y
has generally not been used, since it shows a good positive correlation with the
character X and it also represents a smaller number of chambers than character X
(Drooger, 1952, 1993; Raju,
1974). Also, it is generally more
difficult to determine character Y compared with character X since the first
chamber with two stolons may be difficult to pick in poorly preserved specimens.
) represents the number of chambers in the neorotalid spire
(excluding
P and D) up to, but NOT including the first chamber that gives rise to the fan
of equatorial chambers (Drooger, 1952; Raju,
1974). Character Y
has generally not been used, since it shows a good positive correlation with the
character X and it also represents a smaller number of chambers than character X
(Drooger, 1952, 1993; Raju,
1974). Also, it is generally more
difficult to determine character Y compared with character X since the first
chamber with two stolons may be difficult to pick in poorly preserved specimens.
Character
Z (Fig. 1.A ![]() ) is the number of spiral chambers in the rotalid coil excluding the
first two chambers (P and D), up to and including the largest chamber (Drooger,
1952). The identification of the largest chamber may be difficult (subjective)
in some forms and character Z has not generally been used in the discrimination
of species.
) is the number of spiral chambers in the rotalid coil excluding the
first two chambers (P and D), up to and including the largest chamber (Drooger,
1952). The identification of the largest chamber may be difficult (subjective)
in some forms and character Z has not generally been used in the discrimination
of species.
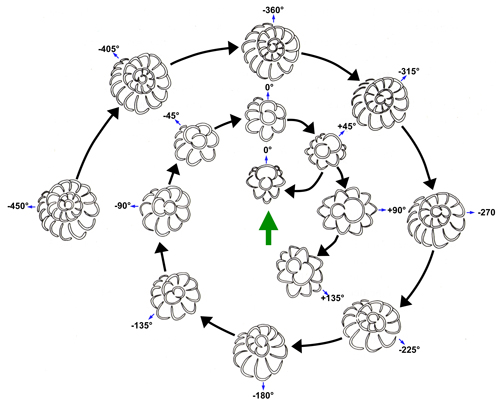
Angle γ
relates the orientation of the medial line through the embryo (centres of P and
D) to the medial line through the fan of chambers forming the test (Drooger,
1952; Fig. 1.B ![]() ). Values of γ range from about -450 to +135 (Fig.
2
). Values of γ range from about -450 to +135 (Fig.
2 ![]() ) and there is a good correlation between Xm
values and γm
values (Raju, 1974; Drooger, 1993). The measurement of γ
has presented problems and Drooger (1993, Fig. 47) introduced a useful
diagram for visualizing different values of γ (redrawn with amendments in Fig.
2
) and there is a good correlation between Xm
values and γm
values (Raju, 1974; Drooger, 1993). The measurement of γ
has presented problems and Drooger (1993, Fig. 47) introduced a useful
diagram for visualizing different values of γ (redrawn with amendments in Fig.
2 ![]() ). Angle
γ is negative for more tightly coiled specimens and
positive for many biserial forms. Unfortunately, some authors (e.g., Boudagher-Fadel & Price,
2010, Fig. 3.B; Novandaru
et al., 2025, Fig. 4.b) have published
figures that show an incorrect method for the measurement of γ and,
unfortunately, their values for γ cannot be trusted.
). Angle
γ is negative for more tightly coiled specimens and
positive for many biserial forms. Unfortunately, some authors (e.g., Boudagher-Fadel & Price,
2010, Fig. 3.B; Novandaru
et al., 2025, Fig. 4.b) have published
figures that show an incorrect method for the measurement of γ and,
unfortunately, their values for γ cannot be trusted.
|
|
Figure 2: Range of variation of γ in different specimens of miogypsinid foraminfers (revised
from Drooger, 1993, Fig. 47). Specimens are arranged with the apical
region orientated towards the top (green arrow). The small blue arrow points
along the embryonic axis towards the deuteroconch. The black arrows show the
evolutionary trend towards less negative γ
values and eventually positive γ values. |
Character
V (Fig. 1.C ![]() ) is a measure of the asymmetry of the two spires of chambers that
emanate from the two PACs and meet in a closing chamber in more advanced ('biserial')
species of Miogypsina (Drooger,
1952). It is calculated as 200-times the ratio of the shorter arc of chambers
(angle α)
divided by the complete arc of chambers (β)
and ranges from 0 (when there is just 1 PAC, i.e., α
= 0) to 100 when the embryo is symmetrical. For species discrimination it is the
mean value of V (Vm) that is important. Character V has been widely
used in defining chronospecies within biserial Miogypsina
populations, yet V values are quite variable among specimens from a single
population (Drooger, 1952, Table 2).
) is a measure of the asymmetry of the two spires of chambers that
emanate from the two PACs and meet in a closing chamber in more advanced ('biserial')
species of Miogypsina (Drooger,
1952). It is calculated as 200-times the ratio of the shorter arc of chambers
(angle α)
divided by the complete arc of chambers (β)
and ranges from 0 (when there is just 1 PAC, i.e., α
= 0) to 100 when the embryo is symmetrical. For species discrimination it is the
mean value of V (Vm) that is important. Character V has been widely
used in defining chronospecies within biserial Miogypsina
populations, yet V values are quite variable among specimens from a single
population (Drooger, 1952, Table 2).
Diameter
Pw (DI) is the width of the protoconch (Pw) measured perpendicular to
the medial line of the embryo (Drooger, 1952). This measurement, by
definition, includes half the thickness of the embryonic wall (Fig.
1.D ![]() ). Values
of X and V show positive correlations with Pw (Raju, 1974; Drooger,
1993).
). Values
of X and V show positive correlations with Pw (Raju, 1974; Drooger,
1993).
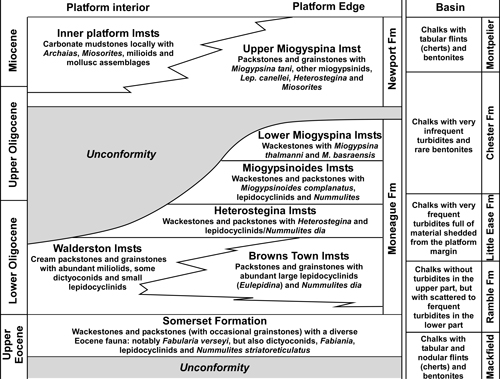
During
the mid Cenozoic, Jamaica was represented by a series of carbonate platforms
surrounded by deep-water troughs (Eva & McFarlane,
1985; Robinson
& Mitchell, 1999; Fig. 3 ![]() ) - a similar set of depositional
environments to those found in the modern-day Bahamas (e.g., Fauquembergue et
al., 2024; Lopez-Gamundim et
al., 2025). Defining lithostratigraphic schemes for the limestones on these
blocks is problematic, because the limestones show relatively few lithological
differences and many can only be distinguished by using biostratigraphy. Three
(Somerset, Moneague, and Newport) formations are accepted here for the Upper
Eocene to mid Miocene succession, with significant unconformities represented by
the bases of the Somerset and Newport formations (Hose & Versey,
1956; Versey in Zans et al.,
1963; Mitchell, 2004, 2013). The Moneague and Newport formations are
divided into a series of 'beds' that are based on their LBF assemblages (Fig. 4
) - a similar set of depositional
environments to those found in the modern-day Bahamas (e.g., Fauquembergue et
al., 2024; Lopez-Gamundim et
al., 2025). Defining lithostratigraphic schemes for the limestones on these
blocks is problematic, because the limestones show relatively few lithological
differences and many can only be distinguished by using biostratigraphy. Three
(Somerset, Moneague, and Newport) formations are accepted here for the Upper
Eocene to mid Miocene succession, with significant unconformities represented by
the bases of the Somerset and Newport formations (Hose & Versey,
1956; Versey in Zans et al.,
1963; Mitchell, 2004, 2013). The Moneague and Newport formations are
divided into a series of 'beds' that are based on their LBF assemblages (Fig. 4 ![]() ). In contrast, the deep-water stratigraphic section can be easily divided into
formations based on lithological criteria, such as, the presence or absence of
flints/cherts and the relative abundance of turbidites (Fig. 4
). In contrast, the deep-water stratigraphic section can be easily divided into
formations based on lithological criteria, such as, the presence or absence of
flints/cherts and the relative abundance of turbidites (Fig. 4 ![]() ). Similar
assemblages of LBF occur allochthonously in the turbidites within the deep-water
succession providing an easy way to correlate between the deep-water and
shallow-water successions. Full lithological and faunal descriptions will be
published elsewhere and they are included here for reference purposes and should
not yet be considered 'formal' or finalized.
). Similar
assemblages of LBF occur allochthonously in the turbidites within the deep-water
succession providing an easy way to correlate between the deep-water and
shallow-water successions. Full lithological and faunal descriptions will be
published elsewhere and they are included here for reference purposes and should
not yet be considered 'formal' or finalized.
Samples were taken from three areas on the Clarendon Block, with the
zonation of the Oligocene to early Miocene based on LBF utilizing the American
Benthic Zonation (ABZ) nomenclature of Mitchell et
al. (2022, 2024). The zonation for the late Oligocene is refined in Table 1
based on the evolution of miogypsinids demonstrated here. Sample locations are
shown in Figures 3 ![]() and 5
and 5 ![]() .
.
|
|
Figure 3: Simplified geological map showing deep-water basins (minor blocks in eastern
and western Jamaica omitted) and the large, shallow-water Clarendon Block in
Oligocene to early Miocene times. Cretaceous rocks (inliers), parish boundaries,
and major roads are shown for reference. The location of the detailed map for
samples around Ulster Spring is shown (yellow box at exaggerated size) together
with other samples locations across Jamaica. |
|
|
Figure 4: Subdivision of part
(uppermost Eocene to mid Miocene) of the shallow-water
White Limestone succession on the Clarendon Block (formations and beds based on
lithology and foraminiferal assemblages) and the deep-water White Limestone
(formations) in the basin. Formal descriptions will be published elsewhere. |
|
|
Figure 5: Detailed map of the area to the west of Ulster Spring (Trelawny) showing
samples (those with miogypsinids in blue). The road transect (samples WL3804 -
WL3810 shows foraminifers in their correct stratigraphic positions according to
the theory of nepionic acceleration (Tan, 1936,
1937; Drooger,
1952) suggesting a low dip of the strata to the east (the limestones are largely
unbedded). |
Table 1. Samples with miogypsinids (arranged by chronospecies) used in this study. All slabs and thins sections are in the Simon F. Mitchell Collection currently stored in collections of the University of the West Indies Geology Museum (UWIGM).
|
Sample |
GPS location |
Details |
|
WL5008 |
Mio. tani, Upper Chester Fm, near mouth of Swift River, Hope Bay, Portland. |
|
|
WL1727 |
Mio. tani, Upper Chester Fm, North Coast Belt, St Mary. |
|
|
WL5548 |
Mio. tani, Basal Newport Fm, SE of Maggotty, northern St Elizabeth. |
|
|
WL3669 |
Mio. tani, Basal Newport Fm, Ulster Spring area, Trelawny. |
|
|
WL119 |
Mio. tani, Basal Newport Fm, road cut on Highway 2000, St Ann. |
|
|
WL3810 |
Mio. 'basraensis', Lower Miogypsina beds, Moneague Fm, Ulster Spring area, Trelawny. |
|
|
WL497 |
Mio. 'basraensis'. Faulted block, Lower Miogypsina beds, Prickly Pole, St Ann. |
|
|
WL3676 |
Mio. 'basraensis', Lower Miogypsina beds, Moneague Fm, Ulster Spring area, Trelawny. |
|
|
WL3809 |
Mio. thalmanni, Lower Miogypsina beds, Moneague Fm, Ulster Spring area, Trelawny. |
|
|
R1118 |
Ms. complanatus, lower Chester Fm, north of Sherwood Content, Trelawny. |
|
|
WL531 |
Ms. complanatus, Miogypsinoides beds, Moenague Formation, Prickly Pole, St Ann. |
|
|
WL3807 |
Ms. complanatus, Miogypsinoides beds, Moneague Fm, Ulster Spring area, Trelawny. |
|
|
WL4791 |
Ms. complanatus, platform lmsts, Moneague Fm, SW of Browns Town, St Ann. |
|
|
WL5772 |
Ms. complanatus, turbidite in chalks, Chester Fm, Rio Bueno, Trelawny. |
Table 2. Revised Oligocene and lower Miocene American Benthic Zones
|
Zone |
Details |
|
ABZ22 |
Spiroclypeus bullbrooki TRZ. In the lower part of the zone, S. bullbrooki occurs with forms of Mio. tani transitional to Mio. 'globulina' (sample WL5008). This level has yielded M1 planktic foraminifers, but it is before the appearance of NN1 nannofossils (Mitchell et al., 2024) indicating a position in the latest Oligocene. The Oligocene/Miocene boundary is therefore placed at the transition from Mio. tani to Mio. 'globulina' in the Americas. |
|
ABZ21D |
Miogypsina tani PRZ. Upper Chattian. |
|
ABZ21C |
Miogypsina gunteri TRZ. Mid Chattian. |
|
ABZ21B |
Miogypsina 'basraensis' TRZ. Mid Chattian. |
|
ABZ21A |
Miogypsina thalmanni TRZ. Lower Chattian. |
|
ABZ20 |
Miogypsinoides complanata TRZ. In Antigua, Ms. complanata occurs in rocks that have yielded a P22 planktic foraminifer and a NN24 to lower NN25 nannoflora assemblage indicating a level at the transition from the Rupelian to the Chattian (Robinson et al., 2017; Mitchell et al., 2024). |
|
ABZ19 |
Heterostegina antillea PRZ. From the FO of H. antillea to the FO of Ms. complanata. Assemblage includes H. antillea, Lep. yurnagunensis, Lep. parvula, Num. dia, and Neorotalia mecatepecensis; Eu undosa, and Eu. favosa in the lower part; Lep. asterodisca in the upper part. Upper Rupelian. |
|
ABZ18 |
Eulepidina undosa PRZ. From the FO of Eu. undosa until the FO of Heterostegina antillea. Assemblage is characterised by Eu. undosa, Eu. favosa, Lep. yurnagunensis, Lep. parvula, Num. dia, and Neo. mecatepecensis. Mid Rupelian. |
|
ABZ17 |
Eulepidina chaperi PRZ. From the LO of H. ocalana to the FO of Eu. undosa. A low diversity assemblage of LBF with Eu. chaperi, Lep. yurnagunensis, Num. dia, and Neo. mecatepecensis. Lower Rupelian. |
|
ABZ16 |
Heterostegina ocalana TRZ. Last Eocene LBF assemblages. Upper Priabonian. |
Miogypsinid
assemblages were collected from four localities in the deep-water White
Limestone (Fig. 3 ![]() ). Two samples with Ms.
complanatus (WL5772, R1118, Figs. 6
). Two samples with Ms.
complanatus (WL5772, R1118, Figs. 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() - 9
- 9 ![]() ) were collected from the lower part of
the Chester Formation in the parish of Trelawny (Table 1). These come from
graded units with abundant LBF (Fig. 4
) were collected from the lower part of
the Chester Formation in the parish of Trelawny (Table 1). These come from
graded units with abundant LBF (Fig. 4 ![]() ) that are interpreted as turbidites. Two
samples (WL1727, WL5008), containing Mio.
tani, were also collected from the upper part of the Chester Formation, one
each from the parishes of St Mary and Portland. Samples from the mid-Chester
Formation generally lack turbidites, so there are no LBF assemblages. The sample
from Portland was collected from the road section extending along the east bank
southwards of the mouth of the Swift River at Hope Bay. This section (Fig. 9
) that are interpreted as turbidites. Two
samples (WL1727, WL5008), containing Mio.
tani, were also collected from the upper part of the Chester Formation, one
each from the parishes of St Mary and Portland. Samples from the mid-Chester
Formation generally lack turbidites, so there are no LBF assemblages. The sample
from Portland was collected from the road section extending along the east bank
southwards of the mouth of the Swift River at Hope Bay. This section (Fig. 9 ![]() ) is
across the Chattian/Aquitanian boundary (Robinson, 2004; Mitchell et
al., 2024). It has yielded Lower Miocene (NN1 and NN2) nannofossil
assemblages and an Oligocene-Lower Miocene (M1) planktonic foraminifer
assemblage (Blow in Robinson, 1969; Robinson,
2004; Mitchell
et al., 2024). The foraminifer sample
contains Mio. tani and Spiroclypeus
bullbrooki that indicates ABZ22 (Mitchell et
al., 2024; Table 2).
) is
across the Chattian/Aquitanian boundary (Robinson, 2004; Mitchell et
al., 2024). It has yielded Lower Miocene (NN1 and NN2) nannofossil
assemblages and an Oligocene-Lower Miocene (M1) planktonic foraminifer
assemblage (Blow in Robinson, 1969; Robinson,
2004; Mitchell
et al., 2024). The foraminifer sample
contains Mio. tani and Spiroclypeus
bullbrooki that indicates ABZ22 (Mitchell et
al., 2024; Table 2).
|
|
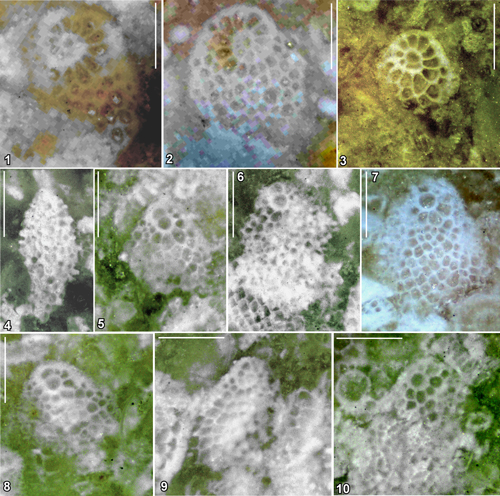
Figure 6: Representative carbonate facies (scans of polished slabs) from Jamaica
containing miogypsinid populations. A (WL5772), turbidite in lower part of the
Chester Formation, Rio Bueno River valley, Trelawny, with Mio. complantus (c) and Heterostegina
antillea (H). B (WL3809), platform
limestone from the Ulster Spring area with Mio. thalmanni (th) and H.
antillea. C (WL3810), platform limestone from the Ulster Spring area with Mio.
'basraensis' (b), H.
antillea, Lepidocyclina asterodisca (La),
H. antillea (H), and Nummulites dia (N).
D (WL5548), platform limestone from the basal Newport Formation SW of northern
Maggotty, St Elizabeth, with Mio. tani
(ta), Lep. asterodisca (La),
and H. antillea (H). |
|
|
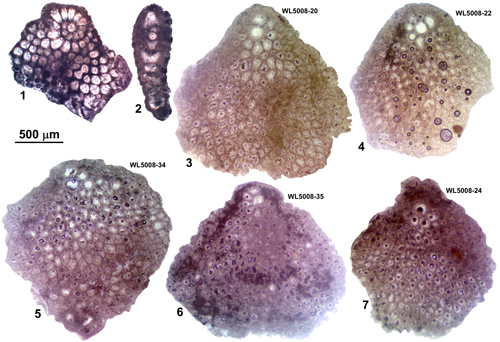
Figure 7: Thin sections of selected miogypsinids from Jamaica. 1 (equatorial), 2
(axial) sections of Miogypsinoides
complanatus, Chester Formation, Sherwood Content). 3-7, equatorial sections
of Miogypsina tani, Chester Formation,
Hope Bay. Scale bar = 500 µm. |
|
|
Figure 8: Photographs of representative specimens of miogypsinids on polished slabs
from Jamaica. 1-2, equatorial sections of Miogypsinoides
companatus, sample WL5772, Chester Formation, Rio Bueno. 3, equatorial
section of Miogypsina basraensis,
sample WL3810, Moneague Formation, Ulster Spring. 4-10, axial (4), and
equatorial (5-10) sections of Miogypsina
tani from sample WL5546, Newport Formation, St Elizabeth. Scale bar = 500 µm. |
|
|
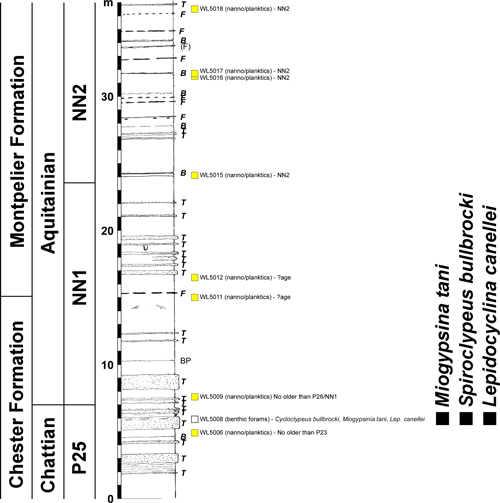
Figure 9: Section across the
Chattian/Aquitanian boundary exposed in the road cutting
along the east bank of the Swift River, south of its mouth, Hope Bay, Portland,
with nannofossil occurrences and foraminifer samples (yellow boxes). The sample
with Mio. tani comes from the upper
part of the Chester Formation in the highest part of the Chattian. T, turbidite;
B, bentonite; BP, bedding plane; F, flint. Partial details have been given by Robinson
(2004) and Mitchell et al. (2024). |
These areas expose extensive swaths of Oligocene and lower Miocene rocks (Hose & Versey, 1956; Versey in Zans et al., 1963; Robinson, 2004; Mitchell, 2004, 2013). On the Manchester plateau, the Moneague Formation is represented by the Walderston beds with at least one intercalation of Browns Town beds. The foraminiferal assemblages of the Walderston beds represent an inner platform facies dominated by miliolids, archaiasids, and Praerhapidionina delicata with sporadic examples of Neorotalia (Robinson & Wright, 1993; Robinson, 2004) with the latter suggesting the Rupelian. The interval with the Browns Town beds contains a LBF assemblage with Eu. undosa and Num. dia without H. antillea (without Heterostegina), indicating ABZ18 (mid Rupelian; Table 2). The Moneague Formation is succeeded unconformably by the Newport Formation, with Versey (in Zans et al., 1963) reported a sparse fauna in the lower part of his Newport Limestones containg Heterostegina, peneroplids, and rare Miogypsina. Stemann (2004) reported a rich coral fauna (32 genera and 64 species) from a section near Albion (Manchester Plateau) at the base of the Newport Limestones. This locality contains Heterostegina antillea and Archaias cf. kirkukensis (Robinson, 2004, Fig. 10.B) and has given a 87Sr/86Sr ration on a Kuphus tube of 0.70823 suggesting, if unaltered, an age of 23.4 to 23.77 Ma [recalibrated from Robinson et al., 2018, based on Geological Timescale 2020 (Raffi et al., 2020)] and attributed to the highest part of the Oligocene (Robinson, 2004; Robinson et al., 2018). According to Robinson et al. (2018), nearby localities have yielded Heterostegina antillea, Miogypsina sp., and small Lepidocyclina sp. Because of its rarity, no suitable samples with populations of Miogypsina have so far been collected from the Manchester Plateau for statistical analyses.
Maggotty
in northern St Elizabeth (Fig. 3 ![]() ) straddles an east-west fault that throws
Eocene rocks (Yellow Limestone and White limestone) to the north against
Oligocene and Miocene rocks to the south. The succession around Maggotty has
yielded good foraminiferal assemblages in the Moneague and Newport formations.
As on the Manchester Plateau, the Moneague Formation consists of Walderston beds
and Browns Town beds, but here the Browns Town beds are thicker and occur in
close proximity, and sometimes immediately below, the Newport Formation. The
Browns Town beds contain an abundant fauna of Eu.
undosa, Num. dia, and Neo.
mecatepecensis, without Heterostegina
antillea, and can be assigned to ABZ18 (Table 2). The basal part of the
overlying Newport Formation contains a very abundant LBF assemblage with H.
antillea, Mio. tani, Lepidocyclina
asterodisca, and Heterostegina
antillea (Fig. 6.D
) straddles an east-west fault that throws
Eocene rocks (Yellow Limestone and White limestone) to the north against
Oligocene and Miocene rocks to the south. The succession around Maggotty has
yielded good foraminiferal assemblages in the Moneague and Newport formations.
As on the Manchester Plateau, the Moneague Formation consists of Walderston beds
and Browns Town beds, but here the Browns Town beds are thicker and occur in
close proximity, and sometimes immediately below, the Newport Formation. The
Browns Town beds contain an abundant fauna of Eu.
undosa, Num. dia, and Neo.
mecatepecensis, without Heterostegina
antillea, and can be assigned to ABZ18 (Table 2). The basal part of the
overlying Newport Formation contains a very abundant LBF assemblage with H.
antillea, Mio. tani, Lepidocyclina
asterodisca, and Heterostegina
antillea (Fig. 6.D ![]() ), and can be assigned to ABZ21D
(Table 2). This is in
good agreement with the Sr-isotopic age from Albion (Manchester) from the base
of the Newport Formation and the sample with advanced Mio. tani from Hope Bay, Portland. The foraminiferal assemblages
from the Maggotty area demonstrate that on this part of the Clarendon Block
rocks containing foraminifers belonging to ABZ19 through ABZ20C have been cut
out by the unconformity at the base of the Newport Formation (Fig.
4
), and can be assigned to ABZ21D
(Table 2). This is in
good agreement with the Sr-isotopic age from Albion (Manchester) from the base
of the Newport Formation and the sample with advanced Mio. tani from Hope Bay, Portland. The foraminiferal assemblages
from the Maggotty area demonstrate that on this part of the Clarendon Block
rocks containing foraminifers belonging to ABZ19 through ABZ20C have been cut
out by the unconformity at the base of the Newport Formation (Fig.
4 ![]() ).
).
In St Ann, on the northern part of the Clarendon Block, it was generally believed that the Miocene succession (Newport Formation) had been removed by erosion caused during the uplift of Jamaica (Hose & Versey, 1956; Versey in Zans et al., 1963; McFarlane, 1977). Robinson et al. (2018) speculated that there might be an outlier of Newport Formation in the Tobolski area based on 87Sr/86Sr dates provided by Land (1991), but mapping in the Tobolski area has not identified anything younger that ABZ19 (Heterostegina beds) or ABZ20 (Miogypsinoides beds) at the present time. Levels high in the Oligocene (ABZ19 through ABZ21B) are, however preserved locally, and levels within the Newport Formation are exposed above the basal-Newport unconformity (with Mio. tani) and in a down-faulted outlier in the Griefield area to the west of Moneague (containing Miogypsina irregularis and Miogypsinita mexicana). The Oligocene miogypsinid assemblages (with Ms. complanata, Mio. 'basraensis', and Mio. tani: Table 2) are considered in this paper, but the Miocene assemblage (with Mio. irregularis and Miog. mexicana) will be discussed elsewhere.
In
this area (Fig. 3 ![]() ), the top of the Moneague Formation and the basal part of the
Newport Formation are preserved in road cuts and yield good assemblages of LBF (Table
1). An excellent road section shows a gently dipping succession of
limestones in the upper part of the Moneague Formation which yields assemblages
showing: ABZ19 (Heterostegina beds, with Neo.
mecatepecensis); ABZ20 (Miogypsinoides
beds, with Ms. complanata); ABZ21A (Miogypsina
beds, with Mio. thalmanni - Fig.
6.B
), the top of the Moneague Formation and the basal part of the
Newport Formation are preserved in road cuts and yield good assemblages of LBF (Table
1). An excellent road section shows a gently dipping succession of
limestones in the upper part of the Moneague Formation which yields assemblages
showing: ABZ19 (Heterostegina beds, with Neo.
mecatepecensis); ABZ20 (Miogypsinoides
beds, with Ms. complanata); ABZ21A (Miogypsina
beds, with Mio. thalmanni - Fig.
6.B ![]() );
and ABZ21B (Miogypsina beds, with Mio. 'basraensis'
- Fig. 6.C
);
and ABZ21B (Miogypsina beds, with Mio. 'basraensis'
- Fig. 6.C ![]() ). Nearby exposures yield Mio.
'basraensis' (WL3676 - from the lower Miogypsina beds of the
Moneagure Formation) and Mio. tani
(WL3669 - from the upper Miogypsina beds of the Newport Formation). Across the
exposures in St Ann and Trelawny, ABZ17 through ABZ21B are present in the
Moneague Formation, overlain unconformably by the Newport Formation (ABZ21D).
Here, the unconformity below the Newport Formation only cuts out ABZ20C (the Mio.
gunteri TRZ).
). Nearby exposures yield Mio.
'basraensis' (WL3676 - from the lower Miogypsina beds of the
Moneagure Formation) and Mio. tani
(WL3669 - from the upper Miogypsina beds of the Newport Formation). Across the
exposures in St Ann and Trelawny, ABZ17 through ABZ21B are present in the
Moneague Formation, overlain unconformably by the Newport Formation (ABZ21D).
Here, the unconformity below the Newport Formation only cuts out ABZ20C (the Mio.
gunteri TRZ).
The study of LBF in the early part of the 20th century in the Americas (as elsewhere) resulted in a proliferation of named miogypsinid species using a typological approach based on small numbers of specimens (Cushman, 1918, 1919; Hodson, 1926; Koch, 1926; Vaughan, 1928a; Gravell, 1933; Nuttall, 1933; Tan, 1936; Cole, 1938; Hanzawa, 1940; Drooger, 1951). While some individual specimens could be related to published 'species', others showed transitional characteristics and were more difficult to place within named 'species'. Using a typological approach, Tan (1936, 1937) introduced the concept of nepionic acceleration in the miogypsinids. His work set out a theoretical evolutionary pathway for the group that showed a progressive reduction in the length of the neorotalid spire over time followed by the development of two PACs.
Drooger (1952) introduced a biometrical approach based on the study of miogypsinid populations from different parts of the Americas. Unfortunately, Drooger (1952) could not independently date his samples and arranged his populations into the hypothetical order based on Tan's (1936, 1937) concept of nepionic acceleration. Subsequently, population studies of miogypsinids were extended into the Neotethys and India (e.g., Drooger, 1954a, 1954b, 1963, 1993; Drooger & Freudenthal, 1964; Drooger & Raju, 1973, 1978; Drooger & Laagland, 1986; amongst others) and have become the standard for the late Oligocene to early Miocene shallow-water benthic zonation using larger foraminifers (Cahuzac & Poignant, 1997). Akers and Drooger (1957) even suggested that the appearance of Miogypsina could be used as a marker for the base of the Miocene for correlation between Europe and the Americas.
In the Americas, palaeontologists rejected the statistical study of populations stating that this was 'very time-consuming and unsatisfactory' (Barker, 1965) or that 'the division is artificial rather than natural' (Cole, 1957, p. 318). Yet Cole would subsequently go on to reduce the number of miogypsinid species in the Americas to five (Cole, 1964) and eventually three (Cole, 1967), recognizing only uniserial forms lacking lateral chambers (Miogypsinoides complanatus), uniserial forms with lateral chambers (Miogypsina gunteri), and biserial forms with lateral chambers (H. antillea). Yet this lumping approach, significantly limited the biostratigraphic value of successive populations (Drooger, 1952, 1963; Akers & Drooger, 1957). Subsequent studies in the Americas have either used excessive lumping, following Cole's 'species' (e.g., Frost & Langenheim, 1974; Robinson, 2004), or typological approaches (e.g., Baumgartner-Mora et al., 2008; Boudagher-Fadel & Price, 2010).
There has also been a progressive splitting of the miogypsinids into genera in different geographical regions based on minor morphological features that have little value when populations are studied. It is worth discussing the development of the classification of the miogypsinids, prior to setting out the simplified formal scheme that is used here.
Barker and Grimsdale (1937) reported the presence of a canal system in the genus Miogypsinoides and postulated that Miogypsinoides had evolved from Rotalia mexicana var. mecatepecensis Nuttall that had a similarly developed canal system. Subsequently, Rotalia mexicana Nuttall was selected by Berm�dez (1952) as the type species for his genus Neorotalia. Some authors (e.g., Cahuzac & Poignant, 1991, 1997, amongst others) have considered that Neorotalia was a junior synonym of Pararotalia Le Calvez and placed Rotalia mexicana and related species in the genus Pararotalia. However, Hottinger et al. (1991) established that, unlike Pararotalia, R. mexicana, possessed an enveloping canal-system (as recorded by Barker & Grimsdale, 1937) and must be attributed generic status. Other authors (e.g., Salmer�n, 1972; Cahuzac & Poignant, 1991, 1997) have recognized that at some levels in Mexico and France specimens of Neorotalia possess one or two 'supplementary equatorial chambers' possibly pre-empting the development of a fan of equatorial chambers that is characteristic of the genus Miogypsinoides. The name Paleomiogypsina boninensis Matsumaru was subsequently introduced by Matsumaru (1996) for similar forms found in Japan, where they were associated with: Neorotalia (cited as Pararotalia) mecatepecensis in his assemblage IV and N. mecatepecensis and Miogypsinella boninensis Matsumaru in his Assemblage V. The generic name Paleomiogypsina is not retained here and is synonymised with Neorotalia since the holotype of P. boniensis came from Assemblage IV without Miogypsinella (Matsumaru, 1996).
The genera Miogypsinoides Hanzawa, 1940, and Miogypsinella Hanzawa, 1940, were established with Miogypsina dehaartii Vlerk, 1924, and Miogypsinella borodinensis Hanzawa, 1940, as type species, respectively. Both lack lateral chambers and only differ in the fact that their early coiled stage shows either planispiral coiling (Miogypsinoides), or low trochospiral coiling (Miogypsinella). Low trochospiral coiling is seen in early forms of Miogypsinoides as well as in the earliest forms of Miogypsina (e.g., specimens of Mio. thalmanni - Drooger, 1952) and there is a progressive change from specimens within successive populations from low trochospiral coiling to planispiral coiling. This is a case of gradational change of characters within populations and it is therefore not logical to split specimens into multiple genera based on such minor gradual changes. As such, the genus Miogypsinella is regarded as a junior synonym of Miogypsinoides (following Loeblich & Tappan, 1988, p. 680).
The phylogenetic development of lateral chamberlets was discussed by Bock (1976). He suggested two methods of formation: 1) as slits originating late in ontogeny between successive laminae added to the sides of the test; and 2) small cavities formed through an association with pillars. The former seems most likely and since it happens in later ontogeny, it would be absent in younger specimens. Subsequently, the genus Postmiogypsinella Sirel & Gedik, 2011 (type species Postmiogypsinella intermedia Sirel & Gedik, 2011), was introduced for specimens with weakly developed lateral chambers. As with Miogypsinella, Postmiogypsinella is considered a transitional form and is therefore regarded as a junior synonym of Miogypsina here.
Boudagher-Fadel and Price (2010, 2013) suggested differences in the wall structure between American (and South African) and Neotethys (Mediterranean)/Indo-Pacific miogypsinids. They stated that "the Mediterranean forms lack one of the typical features of American miogypsinids, namely, strong fissures around the periphery of the test, indicating that they are a distinct, yet parallel, lineage". Yet Bock (1976, p. 6, Fig. 3, Pls. 5 & 8) clearly illustrated that a "marginal fringe" is present in Miogypsinoides bantamensis from France. It is therefore doubtful if this feature can be used to separate miogypsinids from different provinces.
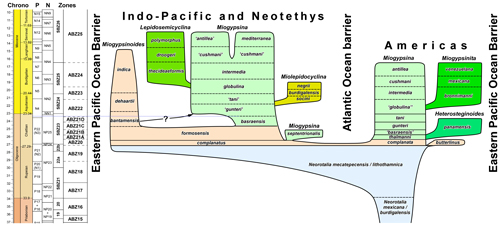
Several different branches extend away from the main Miogypsina lineage, all of which show a relative movement of the neorotalid coil/embryo from a peripheral position towards a more centripetal position within the fan of equatorial chambers. These branches occur at different times within the different provinces and have generally been regarded as subgenera within Miogypsina. The earliest side branch (Heterosteginoides) occurred in the American Province with the development of intercalary chambers within the outer parts of the planispiral neorotalid stage, mimicking genera like Helicostegina. The intercalary chambers begin initially as a single row but increase in numbers to form multiple rows. A second side branch (Miogypsinita) developed in the early Miocene of the American Province, with the development of two spirals developing from each PAC and the fan of equatorial chambers extending around the counter spirals, but the separation of these forms from Miogypsina s.s. is debatable (Drooger, 1993). Miolepidocyclina and Lepidosemicyclina developed as side branches to the main Miogypsina lineage in the Neotethys and Indo-Pacific, respectively, with their own distinct characteristics. The side branches are only dealt with in the evolutionary conclusions in this paper.
A revised classification of Neorotalia and the Miogyspinidae is presented here based on the morphological discussion provided above. At present, it seems likely that Miogypsinoides developed at the same time in all provinces (Americas, Neotethys, and Indo-Pacific). However, the case is probably not the same for Miogypsina, which clearly developed earlier in the Americas than in the Neotethys and Indo-Pacific. At the present time, the name Miogypsina is retained for both lineages, but a separate name (Miogypsinopsis Hanzawa, 1940, being available, but considered a junior synonym of Miogypsina by Loeblich & Tappan, 1988) might be needed for forms in the American Province. Furthermore, Miogypsina species names in all provinces are retained in a purely morphological sense in this paper (i.e., they are 'form chronospecies'). In this paper the 'form chronospecies' names are retained and used in a formal sense (e.g., Miogypsina gunteri) when occurring in the province in which they were named, but in open nomenclature (e.g., Miogypsina 'gunteri') in other provinces. In the future, as more information becomes available, it may be that separate species names will be required for distinct lineages within each province (but his will also require a thorough investigation of all the names that are available in the literature).
Family Calcarinidae Orbigny, 1826
Subfamily Pararotaliinae Reiss, 1963
Genus Neorotalia Berm�dez, 1952
Type species. Rotalia mexicana Nuttall, 1928, from the Eocene of Mexico.
Remarks. If the canal system is taken as a defining characteristic of the Miogypsinidae, then Neorotalia should be transferred to that family and then it can be considered as the route of all subsequent forms. I prefer this course of action.
Family Miogypsinidae Vaughan, 1928b
Genus Miogypsinoides Hanzawa, 1940
Type species. Miogypsina dehaartii Vlerk, 1924, from the Lower Miocene of Larat Island (part of the Tanimbar Islands archipelago) in the province of Maluku, Aru Sea, Indonesia.
Synonyms. Miogypsinella Hanzawa, 1940 (type species Miogypsinella borodinensis Hanzawa, 1940, from the Chattian of Japan); subjective junior synonym.
Remarks. The name Miogypsinoides is used for miogypsinids that have a low trochospiral or planispiral neorotalid stage, have a fan of equatorial chambers and lack lateral chambers. Two species are present in the Americas:
Miogypsinoides
'complanatus' Schlumberger, 1900 (Figs.
7.1-2 ![]() ,
8.1-2
,
8.1-2 ![]() ). The
name is used here (in inverted commas) for American specimens of Miogypsinoides
with Xm between 24, and 17.
). The
name is used here (in inverted commas) for American specimens of Miogypsinoides
with Xm between 24, and 17.
Miogypsinoides butterlinus Salmer�n, 1972. The name is used here for what appears to be a separate branch of Miogypsinoides in the Americas with smaller Xm values (Drooger, 1993). The species has not been reported from Jamaica and is not considered in detail herein.
Genus Miogypsina Sacco, 1893
Type species. Nummulina globulina Michelotti, 1841, by original designation, from the Miocene of Italy.
Synonyms. Flabelliporus Dervieux, 1894 (type species Flabelliporus dilatatus Dervieux, 1894, from the Miocene of Italy). Miogypsinopsis Hanzawa, 1940 (type species Miogypsina (Miogypsina) gunteri Cole, 1938, from the Oligocene of Florida); subjective junior synonym. Miogypsinitella Hanzawa, 1968 (type species Miogypsina (Miogypsina) indonesiensis Tan, 1936, from the Miocene of Java). Tania Matsumaru, 1990 (type species Tania inokosiensis Matsumaru, 1990, from the Miocene of Japan). Postmiogypsinella Sirel & Gedik, 2011 (type species Postmiogypsinella intermedia Sirel & Gedik, 2011, from the Chattian of Turkey).
Remarks. The name Miogypsina is used for miogypsinids with lateral chamberlets. As further work is undertaken, it may be advisable to retain the Neotethys/Indo-Pacific forms within Miogypsina and place the American forms in a separate genus (Miogypsinopsis Hanzawa, 1940). The four branch lines that derived from Miogypsina are given generic status herein. The species of Miogypsina considered here from the Americas are: Mio. thalmanni Drooger, 1952; Mio. 'basraensis' Br�nnimann, 1940; Mio. gunteri Cole, 1938; Mio. tani Drooger, 1952; and Mio. 'irregularis' Michelotti, 1841.
Genus Heterosteginoides Cushman, 1918
Type species. Heterosteginoides panamensis Cushman, 1918, from Oligocene of Panama.
Synonyms. Americogypsina Boudagher-Fadel & Price, 2010 (type species Americogypsina braziliana Boudagher-Fadel & Price, 2010, from the Oligocene of Brazil).
Remarks. This is the earliest branch that develops from Miogypsina in the Americas. Loeblich and Tappan (1988, p. 88, p. 680) considered Heterosteginoides Cushman as a junior synonym of Miolepidocyclina Silvestri. However, Heterosteginoides is a Chattian genus that developed from Miogypsina thalmani in the Americas, whereas Miolepidocyclina is a Burgundian genus that developed from Mio. globuilina in the Neotethys. In my opinion, both genera are valid and developed independently from different miogypsinid ancestors in different palaeogeographic provinces and at different times. The genus is not considered in detail here.
Genus Miogypsinita Drooger, 1952
Type species. Miogypsina mexicana Nuttall, 1933, from the Miocene of Mexico.
Remarks. This is the Miocene branch that develops from Miogypsina in the Americas, but may be difficult to distinguish from Miogypsina (Drooger, 1993). The genus is not considered in detail here.
Genus Lepidosemicyclina Rutten, 1911
Type species. Orbitoides (Lepidosemicyclina) thecideaeformis Rutten, 1911, from the Miocene of Borneo.
Remarks. This is the Miocene branch that develops from Miogypsina in the Indo-Pacific. The genus is not considered in detail here.
Genus Miolepidocyclina Silvestri, 1907
Type species. Orbitoides (Lepidocyclina) burdigalensis G�mbel, 1870, from the Miocene of Italy.
Remarks. This is the Miocene branch that develops from Miogypsina in the Neotethys. The genus is not considered in detail here.
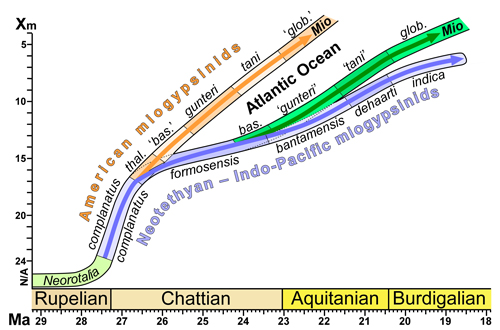
A biometric study of the miogypsinids from the American Province is undertaken here. Both univariate and bivariate statistics are used and comparisons are made, where possible, with other populations from the Americas and populations from the Neotethys and Indo-Pacific. Unfortunately, primary data has not been published for many studies and this makes comparisons difficult. In some cases specimens have been illustrated and can be measured, but this may not include the full range of X, γ, and Pw (etc.) values. I urge that in future studies, all measurements of specimens should be published in full (either is appendices, as supplementary files or in repositories).
Univariant statistical analysis is carried out on three characters here: X, γ, and Pw. Character V is not significant for Oligocene miogypsinids, whereas character Dw (from visual observations) is also unlikely to be of value and was not tested. Therefore, results are provided below for analyses on X, γ, and Pw.
Drooger
(1952, 1993) used mean values of X (Xm) to divide the 'main' Miogypsinoides-Miogypsina
lineage into chronospecies and set limits for each species. The available
populations from Jamaica include sporadic samples from both deep-water and
shallow-water sections and a succession of three samples from the Ulster Spring
Transect. The samples from the Ulster Spring Transect show a progressive
decrease in mean X values (Xm), with populations being assigned to Ms.
complanatus, Mio. thalmanni, and Mio. 'basraensis',
suggesting an evolutionary series (as hypothetically suggested by Drooger,
1952). These and the other populations, from Jamaica are therefore arranged in
order based on their Xm values and assigned to species using Drooger's
(1952, 1993) species limits (Fig. 10 ![]() ). Bar graphs (for continuous data) and
means for γ
and Pw are also plotted on Figure 10 for comparison with dot plots (categorical
data) for X. For γ there is a progressive change towards smaller negative
numbers, whereas for Pw any pattern is less obvious. A dot plot for Drooger's
(1952, 1963) dataset in shown in Figure 11, for comparison.
). Bar graphs (for continuous data) and
means for γ
and Pw are also plotted on Figure 10 for comparison with dot plots (categorical
data) for X. For γ there is a progressive change towards smaller negative
numbers, whereas for Pw any pattern is less obvious. A dot plot for Drooger's
(1952, 1963) dataset in shown in Figure 11, for comparison.
|
|
Figure 10: Dot plots (categorical data) for X and bar graphs (for grouped continuous
data) for γ and Pw for populations from Jamaica,
with means (Xm, γm,
and Pwm). X and γ show progressive changes through populations,
whereas Pw shows little systematic change (although a weak overall trend, with
reversals, to higher values is present). |
|
|
Figure 11: Dot plots (categorical data) for X for populations from Jamaica (upper) and Drooger
(Drooger, 1952, 1963; Akers & Drooger,
1957) (lower)
arranged by Xm with Jamaican dataset tied to ABZ for the mid to late
Oligocene. |
A Kruskal-Wallis rank sum test for multiple independent samples for X on the combined Jamaica-Drooger dataset gave a Kruskal-Wallis chi-squared statistic of 308.4 (26 degrees of freedom), with p = 4.42e-50, showing that some populations were significantly different. Extracted values of p (for the Jamaican, Drooger, and Jamaica-Drooger [Drooger, 1952, 1963; Akers & Drooger, 1957] subsets) from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the combined Jamaican-Drooger (Drooger, 1952, 1963; Akers & Drooger, 1957) dataset are shown in Tables 3 - 4 - 5. These results show high p-values for populations with similar Xm values justifying the use of Xm as a measure for the populations (even if the populations are not normally distributed) and notable statistical differences between Drooger's (1952) chronospecies. Small samples (e.g., WL531) show relative high p-values for both Ms. complanata and Mio. thalmanni (Tables 3, 5), thus demonstrating the problems of trying to identify samples with low numbers of specimens to chronospecies. The results demonstrate that Xm values are very suitable for defining chronospecies in American Oligocene miogypsinids.
Two
notable breaks in the tabulated p-values for populations from Jamaica (Table
3) are apparent: the
first between Ms. complanata and Mio.
thalmanni (excluding WL531 with only 2 specimens) and the second between Mio.
'basraensis' and Mio. tani. This indicates, assuming that the change in populations was due to gradational and
not punctuated evolution, that representatives of all populations were not
sampled. The 'gap' between Ms. complanata
and Mio. thalmanni corresponds to an
influx of platform interior species in which miogypsinids are not present; this
suggests that this 'gap' is due to a facies change. The 'gap' between Mio.
'basraensis' and Mio. tani is
interpreted to represent an unconformity between the underlying Moneague
Formation and the overlying Newport Formation, with the chronospecies Mio.
gunteri missing. As discussed in the geological succession, this
unconformity is clearly demonstrated in the area around Maggotty in northern St.
Elizabeth, where limestones containing ABZ18 foraminiferal assemblages are
overlain by limestones containing ABZ21D foraminiferal assemblages (i.e.,
ABZ19 to ABZ21C are cut out), but only cuts out ABZ21C in the northern part of
the Clarendon Block (Fig. 11 ![]() ).
).
A Kruskal-Wallis rank sum test for multiple independent samples for γ on the Jamaica dataset gave a Kruskal-Wallis chi-squared statistic of 146.6 (13 degrees of freedom), with p = 1.0e-24, showing that some populations are significantly different. Extracted values of p from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the Jamaican dataset are shown in Table 6. The tabulated results show a similar pattern to those for Xm (Table 6) with notable breaks in high p-values between Ms. compalanta and Mio. thalmanni, on the one hand, and Mio. 'basraensis' and Mio. tani, on the other. Clearly, both Xm and γ populations have similar value for discrimination between chronospecies and both should be used. In practice, however, X values are easier to measure than γ values (as is seen in the literature), yet measuring both might provide extra biostratigraphic value.
Table 3. Extracted p-values (arranged by Xm values) for the Jamaican X subset from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the combined Jamaican-Drooger (Drooger, 1952, 1963; Akers & Drooger, 1957) dataset. High probabilities are colour-coded. Samples with low numbers of specimens highlighted. Note the moderately sharp break between Ms. complanata (sample R1118) and Mio. thalmanni (sample WL3809) and the sharper break between Mio. 'basraensis' (sample WL3810) and Mio. tani (sample WL119) suggesting that a continuous suite of populations is not represented.
| Dunn p-values adjusted by the Benjamini-Hochberg FDR method | complanata | thalm. | basraensis | tani | ||||||||||||||
| WL5772 | WL4791 | WL3807 | WL531 | R1118 | WL3809 | WL3676 | WL497 | WL3810 | WL119 | WL3669 | WL5548 | WL1727 | WL5008 | |||||
| Xm | 20.3 | 19.4 | 18.5 | 18.0 | 18.0 | 15.4 | 14.3 | 13.5 | 12.8 | 8.5 | 8.1 | 7.8 | 7.3 | 6.3 | ||||
| Location | Sample | Xm | Xm ID | N | 12 | 7 | 10 | 2 | 30 | 21 | 4 | 11 | 26 | 4 | 18 | 18 | 14 | 9 |
| Chalk | WL5772 | 18.6 | complanata | 12 | 0.87413 | 0.76357 | 0.82514 | 0.54049 | 0.07532 | 0.13784 | 0.01343 | 0.00043 | 0.00019 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL4791 | 19.4 | complanata | 7 | 0.87413 | 0.90383 | 0.90240 | 0.77551 | 0.21251 | 0.22015 | 0.05569 | 0.00830 | 0.00097 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL3807 | 18.5 | complanata | 10 | 0.76357 | 0.90383 | 0.95760 | 0.86779 | 0.21466 | 0.23898 | 0.05024 | 0.00477 | 0.00074 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL531 | 18.0 | complanata | 2 | 0.82514 | 0.90240 | 0.95760 | 0.97933 | 0.59780 | 0.48052 | 0.31279 | 0.19614 | 0.02910 | 0.00777 | 0.00500 | 0.00345 | 0.00174 | |
| Chalk | R1118 | 18.0 | complanata | 30 | 0.54049 | 0.77551 | 0.86779 | 0.97933 | 0.15357 | 0.23898 | 0.02479 | 0.00024 | 0.00032 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL3809 | 15.4 | thalmanni | 21 | 0.07532 | 0.21251 | 0.21466 | 0.59780 | 0.15357 | 0.73099 | 0.35292 | 0.07106 | 0.00713 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL3676 | 14.3 | basraensis | 4 | 0.13784 | 0.22015 | 0.23898 | 0.48052 | 0.23898 | 0.73099 | 0.84626 | 0.61781 | 0.09441 | 0.02063 | 0.01137 | 0.00733 | 0.00383 | |
| Platform | WL497 | 13.5 | basraensis | 11 | 0.01343 | 0.05569 | 0.05024 | 0.31279 | 0.02479 | 0.35292 | 0.84626 | 0.68826 | 0.07040 | 0.00237 | 0.00074 | 0.00053 | 0.00033 | |
| Platform | WL3810 | 12.8 | basraensis | 26 | 0.00043 | 0.00830 | 0.00477 | 0.19614 | 0.00024 | 0.07106 | 0.61781 | 0.68826 | 0.10009 | 0.00122 | 0.00027 | 0.00024 | 0.00020 | |
| Platform | WL119 | 8.5 | tani | 4 | 0.00019 | 0.00097 | 0.00074 | 0.02910 | 0.00032 | 0.00713 | 0.09441 | 0.07040 | 0.10009 | 0.92753 | 0.80426 | 0.67270 | 0.45316 | |
| Platform | WL3669 | 8.1 | tani | 18 | 0.00000 | 0.00000 | 0.00000 | 0.00777 | 0.00000 | 0.00000 | 0.02063 | 0.00237 | 0.00122 | 0.92753 | 0.79415 | 0.58844 | 0.31074 | |
| Platform | WL5548 | 7.8 | tani | 18 | 0.00000 | 0.00000 | 0.00000 | 0.00500 | 0.00000 | 0.00000 | 0.01137 | 0.00074 | 0.00027 | 0.80426 | 0.79415 | 0.78725 | 0.45858 | |
| Chalk | WL1727 | 7.3 | tani | 14 | 0.00000 | 0.00000 | 0.00000 | 0.00345 | 0.00000 | 0.00000 | 0.00733 | 0.00053 | 0.00024 | 0.67270 | 0.58844 | 0.78725 | 0.67270 | |
| Chalk | WL5008 | 6.3 | tani | 9 | 0.00000 | 0.00000 | 0.00000 | 0.00174 | 0.00000 | 0.00000 | 0.00383 | 0.00033 | 0.00020 | 0.45316 | 0.31074 | 0.45858 | 0.67270 | |
| Probabilities | <0.01 | >0.01 | >0.05 | >0.50 | >0.75 | >0.90 | ||||||||||||
Table 4. Extracted p-values (arranged by Xm values) for the Drooger X subset from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the combined Jamaican-Drooger (Drooger, 1952, 1963; Akers & Drooger, 1957) dataset. High probabilities are colour-coded. Note that larger gaps in Xm values (e.g., between Ms. complanata and Mio. 'basraensis', Mio. 'basraensis' and Mio. gunteri and Mio. gunteri and Mio. tani) suggesting that a complete suite of populations has not been sampled.
| Dunn p-values adjusted by the Benjamini-Hochberg FDR method | compl. | thalmanni | basrae. | gunteri | tani | irregularis | |||||||||||
| comp-TT | thal-DR6 | thal-DR4 | thal-LA | bas-TT | gunt-F9 | gunt-F8 | gunt-V | tani-PR | tani-CR | tani-TT | glob-Cu | glob-DR | |||||
| Xm | 23.6 | 15.3 | 15.1 | 15.1 | 13.0 | 11.0 | 10.4 | 10.4 | 8.6 | 7.6 | 7.4 | 7.0 | 5.8 | ||||
| Sample | Xm | Xm ID | N | 5 | 9 | 9 | 16 | 34 | 16 | 16 | 10 | 7 | 11 | 5 | 15 | 6 | |
| comp-TT | 23.6 | complanata | 5 | 0.15639 | 0.13642 | 0.09780 | 0.00662 | 0.00071 | 0.00027 | 0.00067 | 0.00000 | 0.00009 | 0.00002 | 0.00000 | 0.00000 | ||
| thal-DR6 | 15.3 | thalmanni | 9 | 0.15639 | 0.93900 | 0.92274 | 0.23898 | 0.03812 | 0.01564 | 0.03063 | 0.00025 | 0.00453 | 0.00090 | 0.00000 | 0.00007 | ||
| thal-DR4 | 15.1 | thalmanni | 9 | 0.13642 | 0.93900 | 0.98774 | 0.28521 | 0.05015 | 0.02092 | 0.03933 | 0.00035 | 0.00589 | 0.00123 | 0.00000 | 0.00010 | ||
| thal-LA | 15.1 | thalmanni | 16 | 0.09780 | 0.92274 | 0.98774 | 0.19478 | 0.02006 | 0.00635 | 0.01871 | 0.00005 | 0.00214 | 0.00039 | 0.00000 | 0.00002 | ||
| bas-TT | 13.0 | basraensis | 34 | 0.00662 | 0.23898 | 0.28521 | 0.19478 | 0.19478 | 0.08013 | 0.15311 | 0.00069 | 0.02141 | 0.00445 | 0.00000 | 0.00026 | ||
| gunt-F9 | 11.0 | gunteri | 16 | 0.00071 | 0.03812 | 0.05015 | 0.02006 | 0.19478 | 0.76357 | 0.79943 | 0.06399 | 0.24739 | 0.08917 | 0.00139 | 0.01395 | ||
| gunt-F8 | 10.4 | gunteri | 16 | 0.00027 | 0.01564 | 0.02092 | 0.00635 | 0.08013 | 0.76357 | 0.98774 | 0.13653 | 0.40199 | 0.15795 | 0.00531 | 0.03036 | ||
| gunt-V | 10.4 | gunteri | 10 | 0.00067 | 0.03063 | 0.03933 | 0.01871 | 0.15311 | 0.79943 | 0.98774 | 0.17898 | 0.44080 | 0.18916 | 0.01415 | 0.04511 | ||
| tani-PR | 8.6 | tani | 7 | 0.00000 | 0.00025 | 0.00035 | 0.00005 | 0.00069 | 0.06399 | 0.13653 | 0.17898 | 0.74707 | 0.87413 | 0.36401 | 0.43731 | ||
| tani-CR | 7.6 | tani | 11 | 0.00009 | 0.00453 | 0.00589 | 0.00214 | 0.02141 | 0.24739 | 0.40199 | 0.44080 | 0.74707 | 0.65725 | 0.22508 | 0.27819 | ||
| tani-TT | 7.4 | tani | 5 | 0.00002 | 0.00090 | 0.00123 | 0.00039 | 0.00445 | 0.08917 | 0.15795 | 0.18916 | 0.87413 | 0.65725 | 0.60496 | 0.61217 | ||
| irreg-Cu | 7.0 | globulina | 15 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | 0.00139 | 0.00531 | 0.01415 | 0.36401 | 0.22508 | 0.60496 | 0.93224 | ||
| irreg-DR | 5.8 | globulina | 6 | 0.00000 | 0.00007 | 0.00010 | 0.00002 | 0.00026 | 0.01395 | 0.03036 | 0.04511 | 0.43731 | 0.27819 | 0.61217 | 0.93224 | ||
| Probabilities | <0.01 | >0.01 | >0.05 | >0.50 | >0.75 | >0.90 | |||||||||||
Table 5. Extracted p-values (arranged by Xm values) comparing the Jamaica and Drooger X subsets from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the combined Jamaican-Drooger (Drooger, 1952, 1963; Akers & Drooger, 1957) dataset. High probabilities are colour-coded. Samples with low numbers of specimens for the Jamaican dataset highlighted. Note that sample WL5008 has higher p-values when compared against populations of Mio. irregularis than with Mio. tani, suggesting that it may be younger than other populations of Mio. tani.
| Dunn p-values adjusted by the Benjamini-Hochberg FDR method | compl. | thalmanni | basrae. | gunteri | tani | irregularis | |||||||||||
| comp-TT | thal-DR6 | thal-DR4 | thal-LA | bas-TT | gunt-F9 | gunt-F8 | gunt-V | tani-PR | tani-CR | tani-TT | glob-Cu | glob-DR | |||||
| Xm | 23.6 | 15.3 | 15.1 | 15.1 | 13.0 | 11.0 | 10.4 | 10.4 | 8.6 | 7.6 | 7.4 | 7.0 | 5.8 | ||||
| Location | Sample | Xm | Xm ID | N | 5 | 9 | 9 | 16 | 34 | 16 | 16 | 10 | 7 | 11 | 5 | 15 | 6 |
| Chalk | WL5772 | 20.3 | complanata | 12 | 0.84611 | 0.12898 | 0.10314 | 0.05441 | 0.00042 | 0.00003 | 0.00000 | 0.00005 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | 0.00000 |
| Platform | WL4791 | 19.4 | complanata | 7 | 0.76357 | 0.24546 | 0.21665 | 0.16368 | 0.00935 | 0.00083 | 0.00027 | 0.00085 | 0.00000 | 0.00010 | 0.00002 | 0.00000 | 0.00000 |
| Platform | WL3807 | 18.5 | complanata | 10 | 0.65725 | 0.26292 | 0.22888 | 0.16368 | 0.00512 | 0.00035 | 0.00009 | 0.00045 | 0.00000 | 0.00005 | 0.00001 | 0.00000 | 0.00000 |
| Platform | WL531 | 18.0 | complanata | 2 | 0.74707 | 0.58844 | 0.55133 | 0.52566 | 0.21251 | 0.07707 | 0.05015 | 0.05983 | 0.00585 | 0.01762 | 0.00653 | 0.00074 | 0.00153 |
| Chalk | R1118 | 18.0 | complanata | 30 | 0.50574 | 0.24546 | 0.20616 | 0.11006 | 0.00017 | 0.00001 | 0.00000 | 0.00005 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | 0.00000 |
| Platform | WL3809 | 15.4 | thalmanni | 21 | 0.12898 | 0.93784 | 0.86779 | 0.83322 | 0.07929 | 0.00564 | 0.00133 | 0.00644 | 0.00000 | 0.00066 | 0.00011 | 0.00000 | 0.00000 |
| Platform | WL3676 | 14.3 | basraensis | 4 | 0.14528 | 0.79248 | 0.83450 | 0.83450 | 0.67270 | 0.24546 | 0.16394 | 0.19478 | 0.01491 | 0.05524 | 0.01896 | 0.00105 | 0.00370 |
| Platform | WL497 | 13.5 | basraensis | 11 | 0.03316 | 0.51432 | 0.57387 | 0.52530 | 0.76357 | 0.18894 | 0.09544 | 0.14428 | 0.00224 | 0.02517 | 0.00612 | 0.00002 | 0.00055 |
| Platform | WL3810 | 12.8 | basraensis | 26 | 0.00600 | 0.21251 | 0.24946 | 0.17042 | 0.89318 | 0.26546 | 0.13000 | 0.20554 | 0.00167 | 0.03316 | 0.00713 | 0.00000 | 0.00050 |
| Platform | WL119 | 8.5 | tani | 4 | 0.00066 | 0.01871 | 0.02264 | 0.01444 | 0.07907 | 0.36401 | 0.51894 | 0.53451 | 0.78242 | 0.99569 | 0.70425 | 0.32854 | 0.35794 |
| Platform | WL3669 | 8.1 | tani | 18 | 0.00000 | 0.00023 | 0.00033 | 0.00002 | 0.00038 | 0.08815 | 0.19177 | 0.24546 | 0.77092 | 0.91472 | 0.67454 | 0.15311 | 0.24546 |
| Platform | WL5548 | 7.8 | tani | 18 | 0.00000 | 0.00007 | 0.00010 | 0.00000 | 0.00007 | 0.03885 | 0.09979 | 0.15311 | 0.94104 | 0.76357 | 0.82514 | 0.24546 | 0.35230 |
| Chalk | WL1727 | 7.3 | tani | 14 | 0.00000 | 0.00005 | 0.00007 | 0.00000 | 0.00007 | 0.02457 | 0.06366 | 0.10051 | 0.86574 | 0.59780 | 0.98724 | 0.46785 | 0.52566 |
| Chalk | WL5008 | 6.3 | tani | 9 | 0.00000 | 0.00003 | 0.00005 | 0.00000 | 0.00007 | 0.01223 | 0.03036 | 0.05015 | 0.55706 | 0.35759 | 0.76059 | 0.86779 | 0.83450 |
| Probabilities | <0.01 | >0.01 | >0.05 | >0.50 | >0.75 | >0.90 | |||||||||||
Table 6. Extracted p-values (arranged by Xm values) for the Jamaica γ dataset from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method). High probabilities are colour-coded. Samples with low numbers of specimens for the Jamaican dataset highlighted.
| Dunn p-values adjusted by the Benjamini-Hochberg FDR method | complanata | thal. | basraensis | tani | ||||||||||||||
| WL4791 | WL5772 | WL3807 | WL531 | R1118 | WL3809 | WL3676 | WL497 | WL3810 | WL119 | WL3669 | WL5548 | WL1727 | WL5008 | |||||
| Xm | 18.6 | 19.4 | 18.5 | 18.0 | 18.0 | 15.4 | 14.3 | 13.5 | 12.8 | 8.5 | 8.1 | 7.8 | 7.3 | 6.3 | ||||
| Location | Sample | Xm | Xm ID | N | 7 | 5 | 10 | 2 | 30 | 21 | 4 | 11 | 26 | 4 | 18 | 18 | 14 | 9 |
| Platform | WL4791 | 18.6 | complanata | 7 | 0.81609 | 0.88594 | 0.84212 | 0.97876 | 0.31394 | 0.31929 | 0.08808 | 0.03830 | 0.00280 | 0.00031 | 0.00001 | 0.00001 | 0.00001 | |
| Chalk | WL5772 | 19.4 | complanata | 5 | 0.81609 | 0.85719 | 0.99377 | 0.80834 | 0.26009 | 0.25017 | 0.09712 | 0.06601 | 0.00427 | 0.00273 | 0.00035 | 0.00029 | 0.00028 | |
| Platform | WL3807 | 18.5 | complanata | 10 | 0.88594 | 0.85719 | 0.88594 | 0.88594 | 0.16280 | 0.22504 | 0.03331 | 0.00806 | 0.00105 | 0.00001 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL531 | 18.0 | complanata | 2 | 0.84212 | 0.99377 | 0.88594 | 0.84212 | 0.36171 | 0.31929 | 0.17546 | 0.13765 | 0.01248 | 0.01316 | 0.00284 | 0.00242 | 0.00195 | |
| Chalk | R1118 | 18.0 | complanata | 30 | 0.97876 | 0.80834 | 0.88594 | 0.84212 | 0.09383 | 0.22476 | 0.01367 | 0.00061 | 0.00058 | 0.00000 | 0.00000 | 0.00000 | 0.00000 | |
| Platform | WL3809 | 15.4 | thalmanni | 21 | 0.31394 | 0.26009 | 0.16280 | 0.36171 | 0.09383 | 0.82514 | 0.34195 | 0.17546 | 0.01124 | 0.00041 | 0.00000 | 0.00000 | 0.00001 | |
| Platform | WL3676 | 14.3 | basraensis | 4 | 0.31929 | 0.25017 | 0.22504 | 0.31929 | 0.22476 | 0.82514 | 0.84212 | 0.79875 | 0.08028 | 0.09996 | 0.01999 | 0.01492 | 0.01252 | |
| Platform | WL497 | 13.5 | basraensis | 11 | 0.08808 | 0.09712 | 0.03331 | 0.17546 | 0.01367 | 0.34195 | 0.84212 | 0.92204 | 0.07479 | 0.05543 | 0.00320 | 0.00280 | 0.00290 | |
| Platform | WL3810 | 12.8 | basraensis | 26 | 0.03830 | 0.06601 | 0.00806 | 0.13765 | 0.00061 | 0.17546 | 0.79875 | 0.92204 | 0.06601 | 0.02077 | 0.00041 | 0.00045 | 0.00085 | |
| Platform | WL119 | 8.5 | tani | 4 | 0.00280 | 0.00427 | 0.00105 | 0.01248 | 0.00058 | 0.01124 | 0.08028 | 0.07479 | 0.06601 | 0.54765 | 0.90533 | 0.99883 | 0.88594 | |
| Platform | WL3669 | 8.1 | tani | 18 | 0.00031 | 0.00273 | 0.00001 | 0.01316 | 0.00000 | 0.00041 | 0.09996 | 0.05543 | 0.02077 | 0.54765 | 0.31929 | 0.24300 | 0.18421 | |
| Platform | WL5548 | 7.8 | tani | 18 | 0.00001 | 0.00035 | 0.00000 | 0.00284 | 0.00000 | 0.00000 | 0.01999 | 0.00320 | 0.00041 | 0.90533 | 0.31929 | 0.85719 | 0.68688 | |
| Chalk | WL1727 | 7.3 | tani | 14 | 0.00001 | 0.00029 | 0.00000 | 0.00242 | 0.00000 | 0.00000 | 0.01492 | 0.00280 | 0.00045 | 0.99883 | 0.24300 | 0.85719 | 0.84212 | |
| Chalk | WL5008 | 6.3 | tani | 9 | 0.00001 | 0.00028 | 0.00000 | 0.00195 | 0.00000 | 0.00001 | 0.01252 | 0.00290 | 0.00085 | 0.88594 | 0.18421 | 0.68688 | 0.84212 | |
| Probabilities | <0.01 | >0.01 | >0.05 | >0.50 | >0.75 | >0.90 | ||||||||||||
A Kruskal-Wallis rank sum test for multiple independent samples for Pw on the Jamaica dataset gave a Kruskal-Wallis chi-squared statistic of 63.0 (13 degrees of freedom) with p = 1.51e-08 (but note the orders of magnitude difference the chi-squared statistic between X, γ, and Pw), showing that some populations are significantly different. Extracted values of p from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method) for the Jamaican dataset are shown in Table 7. The tabulated results show much lower p-values for comparisons between pairs of populations, with some populations (e.g., WL3810) showing low p-values for all populations. This suggests that Pw values have low value in discriminating species in Miogypsinoides and early Miogypsina populations in the Americas. It has been established (e.g., Nigam & Rao, 1987; Bryan, 1995; Yu et al., 2016) that Pw is dependent on various environmental conditions (e.g., water depth) and it clearly is less suitable to separate chronospecies in Oligocene miogypsinids.
Table 7. Extracted p-values (arranged by Xm values) for the Jamaica Pw dataset from a pairwise Dunn post-hoc test (adjusted by the Benjamini-Hochberg FDR method). High probabilities are colour-coded. Samples with low numbers of specimens for the Jamaican dataset highlighted. Probabilities are considerably lower than for corresponding tests for X and γ suggesting that Pw is not a good discriminator at the species level and that other factors may be influencing Pw.
| Dunn p-values adjusted by the Benjamini-Hochberg FDR method | complanata | thal. | basraensis | tani | ||||||||||||||
| WL4791 | WL5772 | WL3807 | WL531 | R1118 | WL3809 | WL3676 | WL497 | WL3810 | WL119 | WL3669 | WL5548 | WL1727 | WL5008 | |||||
| Xm | 18.6 | 19.4 | 18.5 | 18.0 | 18.0 | 15.4 | 14.3 | 13.5 | 12.8 | 8.5 | 8.1 | 7.8 | 7.3 | 6.3 | ||||
| Locality | Sample | Xm | Xm ID | N | 7 | 5 | 10 | 2 | 30 | 21 | 4 | 11 | 26 | 4 | 18 | 18 | 14 | 9 |
| Platform | WL4791 | 18.6 | complanata | 7 | 0.86325 | 0.86404 | 0.57515 | 0.15175 | 0.24326 | 0.65818 | 0.32255 | 0.06294 | 0.05274 | 0.02471 | 0.00361 | 0.01313 | 0.00009 | |
| Chalk | WL5772 | 19.4 | complanata | 5 | 0.86325 | 0.94362 | 0.53412 | 0.40513 | 0.50464 | 0.80387 | 0.55471 | 0.23516 | 0.10575 | 0.12816 | 0.05324 | 0.08385 | 0.00360 | |
| Platform | WL3807 | 18.5 | complanata | 10 | 0.86404 | 0.94362 | 0.49611 | 0.12950 | 0.23725 | 0.71216 | 0.33563 | 0.04484 | 0.05274 | 0.01317 | 0.00099 | 0.00474 | 0.00001 | |
| Platform | WL531 | 18.0 | complanata | 2 | 0.57515 | 0.53412 | 0.49611 | 0.12950 | 0.16002 | 0.38889 | 0.19376 | 0.07450 | 0.04044 | 0.04044 | 0.01628 | 0.02471 | 0.00099 | |
| Chalk | R1118 | 18.0 | complanata | 30 | 0.15175 | 0.40513 | 0.12950 | 0.12950 | 0.76902 | 0.53984 | 0.74524 | 0.49020 | 0.18147 | 0.15175 | 0.02310 | 0.07648 | 0.00022 | |
| Platform | WL3809 | 15.4 | thalmanni | 21 | 0.24326 | 0.50464 | 0.23725 | 0.16002 | 0.76902 | 0.66201 | 0.92924 | 0.36239 | 0.15175 | 0.12816 | 0.02233 | 0.06294 | 0.00022 | |
| Platform | WL3676 | 14.3 | basraensis | 4 | 0.65818 | 0.80387 | 0.71216 | 0.38889 | 0.53984 | 0.66201 | 0.71216 | 0.42072 | 0.12950 | 0.15175 | 0.06294 | 0.10575 | 0.00360 | |
| Platform | WL497 | 13.5 | basraensis | 11 | 0.32255 | 0.55471 | 0.33563 | 0.19376 | 0.74524 | 0.92924 | 0.71216 | 0.39537 | 0.15175 | 0.15175 | 0.04044 | 0.08813 | 0.00089 | |
| Platform | WL3810 | 12.8 | basraensis | 26 | 0.06294 | 0.23516 | 0.04484 | 0.07450 | 0.49020 | 0.36239 | 0.32072 | 0.39537 | 0.32072 | 0.49611 | 0.12816 | 0.27498 | 0.00245 | |
| Platform | WL119 | 8.5 | tani | 4 | 0.05274 | 0.10575 | 0.05274 | 0.04044 | 0.18147 | 0.15175 | 0.12950 | 0.15175 | 0.32072 | 0.50464 | 0.71216 | 0.63373 | 0.60114 | |
| Platform | WL3669 | 8.1 | tani | 18 | 0.02471 | 0.12816 | 0.01317 | 0.04044 | 0.15175 | 0.12816 | 0.15175 | 0.15175 | 0.49611 | 0.50464 | 0.49611 | 0.71216 | 0.02471 | |
| Platform | WL5548 | 7.8 | tani | 18 | 0.00361 | 0.05324 | 0.00099 | 0.01628 | 0.02310 | 0.02233 | 0.06294 | 0.04044 | 0.12816 | 0.71216 | 0.49611 | 0.76284 | 0.08591 | |
| Chalk | WL1727 | 7.3 | tani | 14 | 0.01313 | 0.08385 | 0.00474 | 0.02471 | 0.07648 | 0.06294 | 0.10575 | 0.08813 | 0.27498 | 0.63373 | 0.71216 | 0.76284 | 0.06294 | |
| Chalk | WL5008 | 6.3 | tani | 9 | 0.00009 | 0.00360 | 0.00001 | 0.00099 | 0.00022 | 0.00022 | 0.00360 | 0.00089 | 0.00245 | 0.60114 | 0.02471 | 0.08591 | 0.06294 | |
| Probabilities | <0.01 | >0.01 | >0.05 | >0.50 | >0.75 | >0.90 | ||||||||||||
In this section the relationship between X versus γ and X versus Pw will be explored. This will include analyses of the Jamaican dataset (where complete suites of character values are available) and the global (Americas, Neotethys, and Indo-Pacific) dataset where only mean values are available.
Some scatter plots for X versus γ have previously been published (e.g., Raju, 1974; Drooger, 1993), but have largely presented mean values (Xm and γm) rather than plots for populations. Understanding the fields in which values of X and γ for each population plot is clearly desirable before plotting mean values for multiple populations. Herein, I will start by plotting fields and then look at the distribution patterns of means and ranges for each population. Finally I will compare with published populations (largely mean values).
A scatter
plot for X versus γ
for populations of Oligocene miogypsinids from the Americas (Fig.
12 ![]() ) shows a
strong correlation between X and γ. The fields overlap significantly (as expected) and
show a progressive evolution from high values of X and strongly negative values
of γ to lower values of X and less negative/neutral values
of γ. This reflects the trends in the univariate analyses
for both X and γ. A scatterplot of Xm and γm
values for the American, Neotethys and Indo-Pacific populations shows similar
relations (Fig. 13
) shows a
strong correlation between X and γ. The fields overlap significantly (as expected) and
show a progressive evolution from high values of X and strongly negative values
of γ to lower values of X and less negative/neutral values
of γ. This reflects the trends in the univariate analyses
for both X and γ. A scatterplot of Xm and γm
values for the American, Neotethys and Indo-Pacific populations shows similar
relations (Fig. 13 ![]() ), indicating the relationship between these variables is
similar in all three provinces (although there is a departure of γ
toward more negative values for Xm = 11 to 18 for the Indo-Pacific
Province). Since the number of chambers in the spire should be related to the
orientation of the embryonic axis this is not a surprising result.
), indicating the relationship between these variables is
similar in all three provinces (although there is a departure of γ
toward more negative values for Xm = 11 to 18 for the Indo-Pacific
Province). Since the number of chambers in the spire should be related to the
orientation of the embryonic axis this is not a surprising result.
|
|
Figure 12: Scatter plot for X against γ
for populations of uniserial miogypsinids from the Americas. Note the strong
correlation (linear trend). Jamaican samples named on the right, other samples
on the left. |
|
|
Figure 13: Scatter plot for Xm against γm
for populations of uniserial miogypsinids from the Americas, Neotethys, and
Indo-Pacific. Note the strong correlation (linear trend) showing that all
provinces show a similar type of development with regard to Xm versus
γm
(but not necessarily at the same time). Note however that Indo-Pacific values
plot to the left (lower γm
values) compared to Tethys and American Xm values between around 20
and 11. Data from: Drooger, 1954a,
1954b, 1963; Drooger et
al., 1955; Drooger & Freudenthal,
1964; Ferrero, 1965, 1987; Soediono,
1969; Cerutti,
1973; Mulder, 1975; Bock,
1976, 1977; Sch�ttenhelm,
1976; Schiavinotto, 1979,
1985; Delicat
& Schiavinotto, 1985; Wildenborg,
1991; Ferrero et
al., 1994; �zcan et al.,
2010. |
Figure 14 shows a scatterplot for X versus Pw for the Jamaican dataset. Although there is a gradual increase in Pw values for increasing X values, the trend cannot be used to separate chronospecies.
|
|
Figure 14: Scatterplot
of X versus Pw (in µm) for Jamaican populations of uniserial miogypsinids. Pw
shows only a very gradual increase and cannot be used to separate chronospecies.
Note that the youngest sample (WL5008), however, has notably higher values. |
|
|
Figure 15: Scatterplot
of Xm versus Pwm for Jamaican dataset and selected other
populations (Drooger, 1952, 1993; Raju,
1974). This scatterplot
considers only morphology and not age of populations. Note the broad
Miogypsina field, with populations from the Americas occupying the whole
width of the field, yet those from Jamaica (a single carbonate platform) only
occupy the right-hand side of the field). Similar restrictions to parts of the
fields for Miogypsina
and Miogypsinoides are shown for populations from different
geographical areas (e.g., India and Neotethys). |
Scatter
plots for selected global populations of Xm versus Pwm
show broad fields for Miogypsina and Miogypsinoides
with populations from some geographical areas only occupying parts of each field
(Fig. 15 ![]() ). The field for Miogypsina
for the Americas is wide and occupies most of the field for all Miogypsina populations
(Fig. 15
). The field for Miogypsina
for the Americas is wide and occupies most of the field for all Miogypsina populations
(Fig. 15 ![]() ), yet the populations from Jamaica
only occupy part of the overall field. Similarly, European populations (Eu)
occupy the right-hand side of the Miogypsina
field, whereas Indian populations (In) occupy the left-hand side of the field.
The populations in the Miogypsinoides
field generally have larger Pwm values and European (Eu) and Indian
(In) populations occupy different parts of the field. Other Miogypsinoides
populations (g - Guinea; e - Egypt) occupy parts of the Miogypsina
field, whereas Miogypsina septentrinalis
(d - from Germany) forms a small field with larger Pwm values and
there are other populations that represent outliers. This suggests that
populations with local Pwm values developed on a particular carbonate
platform (e.g., the Miogypsinoides-Miogypsina lineage on the Clarendon Block for
Jamaica) or within a
restricted geographical area. Consequently, the use of Pwm to define
chronospecies would seem to be unwise.
), yet the populations from Jamaica
only occupy part of the overall field. Similarly, European populations (Eu)
occupy the right-hand side of the Miogypsina
field, whereas Indian populations (In) occupy the left-hand side of the field.
The populations in the Miogypsinoides
field generally have larger Pwm values and European (Eu) and Indian
(In) populations occupy different parts of the field. Other Miogypsinoides
populations (g - Guinea; e - Egypt) occupy parts of the Miogypsina
field, whereas Miogypsina septentrinalis
(d - from Germany) forms a small field with larger Pwm values and
there are other populations that represent outliers. This suggests that
populations with local Pwm values developed on a particular carbonate
platform (e.g., the Miogypsinoides-Miogypsina lineage on the Clarendon Block for
Jamaica) or within a
restricted geographical area. Consequently, the use of Pwm to define
chronospecies would seem to be unwise.
Specimens
of Neorotalia in the Rupelian of the
Americas (Salmer�n, 1972), northern Europe (Cahuzac & Poignant,
1991, 1997), and Japan (Matsumaru,
1990) occasionally develop one or two
equatorial chambers indicating that a few chambers in the neorotalid spire
sometimes developed two stolons. The development of a complete fan of equatorial
chambers (the transition from Neorotalia
to Miogypsinoides) can be placed at
about the boundary between calcareous nannoplankton zones NP22 and NP23 in all
areas (e.g., Cahuzac & Poignant,
1997; Robinson et
al., 2017; Lunt & Luan,
2022; Mitchell et al., 2024). The apparently synchronous development of Miogypsinoides
from Neorotalia in all provinces (Americas, Neotethys, Indo-Pacific)
suggests gene flow at this time between all miogypsinid populations. The
transition between Miogypsinoides and Miogypsina
(the acquisition of lateral chamberlets) occurred sometime during zone NP25,
however, the long duration of this zone makes it impossible to judge if this was
synchronous across all the provinces. The fact that Miogypsina
rapidly evolved in the Americas, but saw slower evolution in the Neotethys and
Indo-Pacific, almost certainly suggest that Miogypsina
developed earlier in the Americas than in the other provinces (Figs.
16 ![]() - 17
- 17 ![]() ). If
this is the case, it would suggest that gene flow had become restricted between
the Americas and the Neotethys with the growing width of the Atlantic becoming a
physical barrier to migration. Perhaps the development of Mio.
septentrionalis in northern Europe was related to the final gene flow across
the Atlantic, but that the main evolutionary development of Miogypsina
in the Neotethys and Indo-Pacific provinces was delayed. Certainly by the mid
Chattian, the evolutionary development of Miogypsina
was well advanced in the American province, but had only just begun in the
Neotethys and Indo-Pacific provinces (Figs.
16
). If
this is the case, it would suggest that gene flow had become restricted between
the Americas and the Neotethys with the growing width of the Atlantic becoming a
physical barrier to migration. Perhaps the development of Mio.
septentrionalis in northern Europe was related to the final gene flow across
the Atlantic, but that the main evolutionary development of Miogypsina
in the Neotethys and Indo-Pacific provinces was delayed. Certainly by the mid
Chattian, the evolutionary development of Miogypsina
was well advanced in the American province, but had only just begun in the
Neotethys and Indo-Pacific provinces (Figs.
16 ![]() - 17
- 17 ![]() ).
).
|
|
Figure 16: Provisional evolution of the
Miogyspinids. The evolution of Miogypsina
in the Indo-Pacific and Neotethys is split by the emergence of a land barrier in
the mid Miocene. Minor offshoots and South African miogypsinids not included. |
|
|
Figure 17: Comparison of the evolutionary changes for uniserial miogypsinids between
the Americas and the Neotethys/Indo-Pacific provinces plotted in Xm
vs. time (Ma) space. Note the distinct time offset (2 to 2.5 Ma) for the 'same'
chronospecies (e.g., gunteri
and tani) between provinces. |
The base
of the Miocene (using the base of NN1 as a proxy) is the next obvious time to
compare the evolutionary development of miogyspinids. In the Americas, the base
of the Miocene corresponds to the transition from Mio.
tani to Mio. 'globulina' (Figs.
16 ![]() - 17
- 17 ![]() ). In contrast, in the Neotethys and
Indo-Pacific provinces the base of the Miocene corresponds to the transition
from Ms. formosensis or Mio.
basraensis to Mio. 'gunteri' (Figs.
16
). In contrast, in the Neotethys and
Indo-Pacific provinces the base of the Miocene corresponds to the transition
from Ms. formosensis or Mio.
basraensis to Mio. 'gunteri' (Figs.
16 ![]() - 17
- 17 ![]() ). Thus, the evolution of the Miogypsina
lineage in the Neotethys and Indo-Pacific was 'delayed' by two chronospecies
(some 2.5 million years) in comparison to the Americas by the start of the
Miocene (Fig. 17
). Thus, the evolution of the Miogypsina
lineage in the Neotethys and Indo-Pacific was 'delayed' by two chronospecies
(some 2.5 million years) in comparison to the Americas by the start of the
Miocene (Fig. 17 ![]() ). Thus, Miogypsina
may potentially be polyphyletic, but more studies of the transition between Miogypsinoides and Miogypsina are
needed to establish this. In the Americas Miogypsinoides disappears, whereas in the Indo-Pacific and Neotethys
a Miogypsinoides lineage persists into
the mid Miocene (Lunt & Luan,
1922; Fig.
16
). Thus, Miogypsina
may potentially be polyphyletic, but more studies of the transition between Miogypsinoides and Miogypsina are
needed to establish this. In the Americas Miogypsinoides disappears, whereas in the Indo-Pacific and Neotethys
a Miogypsinoides lineage persists into
the mid Miocene (Lunt & Luan,
1922; Fig.
16 ![]() ). Short-lived
offshoots occurred throughout the evolution of the miogypsinids (Fig.
16
). Short-lived
offshoots occurred throughout the evolution of the miogypsinids (Fig.
16 ![]() ); I
include in these: Miogypsinoides
butterlinus, Miogypsina
septentrionalis, and four genera (Lepidosemicyclina,
Miolepidocyclina, Heterosteginoides,
and Miogypsinita).
); I
include in these: Miogypsinoides
butterlinus, Miogypsina
septentrionalis, and four genera (Lepidosemicyclina,
Miolepidocyclina, Heterosteginoides,
and Miogypsinita).
A detailed study of the miogypsinids from the Oligocene of Jamaica demonstrates that Drooger's (1952) population approach is valid and that typological-based approaches are of limited use and should not be used. The evolution of Miogypsinoides from Neorotalia seems to be synchronous across all provinces, yet the subsequent development of Miogypsina is probably diachronous, with the American forms showing a more-advanced development than coeval forms from the Neotethys and Indo-Pacific. For the present, the chronospecies names are retained, but with further work separate chronospecies names may well be needed for the Americas and the Neotethys/Indo-Pacific. The base of the Miocene corresponds to the appearance of Mio. 'globulina' in the Americas, but to the appearance of Mio. 'gunteri' in the Neotethys/Indo-Pacific. This indicates that by this time, the miogypsinids of the America were two chronospecies more advanced that their compatriots in the Neotethys/Indo-Pacific. The miogypsinids can be used for detailed biostratigraphy within each province, but cannot be used for correlation between the Americans and the Neotethys/Indo-Pacific. This research demonstrates that LBF lineages can become separated over time and can show different rates of evolution in different, isolated populations.
This work forms part of the study on LBF supported by the prestigious W. Storrs Cole Memorial Research Award managed by the Geological Society of America. This funding is greatly appreciated.
I thank the late Donovan Blissett for helping collect samples with
miogypsinids from Jamaica. Sample R1118 was collected jointly with Natalie Robinson
who also provided a thin section. Mark Jiang analysed the nannofossils
from Hope Bay as part of our study erecting the larger benthic foraminiferal
zonation for the Cenozoic of the Americas (Mitchell et
al., 2024). I thank Michael Hesemann of the Foraminifera.eu Lab
(Hamburg, Germany) for allowing the use of the figure of Miogypsinoides shown in
Figure 1 ![]() (the illustration is from https://foraminifera.eu/contact.html).
Many thanks to Edward (Ted) Robinson for comments on the original
manuscript. Many thanks to David Smith (UWI) and Ian Boxill (UWI)
for reviewing the use of statistics in this paper. I thank the reviewers,
Lorenzo Consorti and one anonymous, for their comments that helped
improve the original paper. I thank the journal editor, Alberto Collareta,
for his swift handing of this paper.
(the illustration is from https://foraminifera.eu/contact.html).
Many thanks to Edward (Ted) Robinson for comments on the original
manuscript. Many thanks to David Smith (UWI) and Ian Boxill (UWI)
for reviewing the use of statistics in this paper. I thank the reviewers,
Lorenzo Consorti and one anonymous, for their comments that helped
improve the original paper. I thank the journal editor, Alberto Collareta,
for his swift handing of this paper.
Akers W.H. & Drooger C.W. (1957).- Miogypsinids, planktonic Foraminifera, and Gulf Coast Oligocene-Miocene correlations.- AAPG Bulletin, Tulsa - OK, vol. 41, no. 4, p. 656-674.
Barker R.W. (1965).- Notes on Miogypsinidae in the Gulf of Mexico region. In: Dr. D.N. Wadia commemorative volume.- Mining, Geological & Metallurgical Institute of India, Kolkata, p. 306-342 [Not seen].
Barker R.W. & Grimsdale T.F. (1937).- XIV.- Studies of Mexican fossil Foraminifera.- Annals and Magazine of Natural History (10th Serie), vol. XIX, no. 110, p. 161-178.
Baumgartner-Mora C., Baumgartner P.O. & Tschudin P. (2008).- Late Oligocene larger foraminifera from Nosara, Nicoya Pen�nsula (Costa Rica) and Windward, Carriacou (Lesser Antilles), calibrated by 87Sr/86Sr isotope stratigraphy.- Revista Geol�gica de Am�rica Central, San Jos�, vol. 38, p. 33-52. DOI: 10.15517/rgac.v0i38.4215
Benjamini Y. & Hochberg Y. (1995).- Controlling the false discovery rate: A practical and powerful approach to multiple testing.- Journal of the Royal Statistical Society (Series B), vol. 57, no. 1, p. 289-300. DOI: 10.1111/j.2517-6161.1995.tb02031.x
Berm�dez P.J. (1952).- Estudio sistematico de los Foraminiferos rotaliformes.- Rep�blica de Venezuela, Ministerio de Minas e Hidrocarburos, Boletin de Geologia, Caracas, vol. 2, no. 4, p. 1-230.
Bock J.F. de (1976).- Studies on some Miogypsinoides - Miogypsina s.s. associations with special reference to morphological features.- Scripta Geologica, Leiden, vol. 36, p. 1-135. URL: https://repository.naturalis.nl/pub/317461
Bock J.F. de (1977).- Some remarks on Miogypsina (Miogypsina) socini Drooger, 1954 (Foraminifera) from Northern ltaly.- Scripta Geologica, Leiden, vol. 40, p. 1-35. URL: https://repository.naturalis.nl/pub/317497
BouDagher-Fadel M.K. & Price G.D. (2010).- American Miogypsinidae: An analysis of their phylogeny and biostratigraphy.- Micropaleontology, Flushing - NY, vol. 56, no. 6, p. 567-586.
Boudagher-Fadel M.K. & Price G.D. (2013).- The phylogenetic and palaeogeographic evolution of the miogypsinid larger benthic Foraminifera.- Journal of the Geological Society, London, vol. 170, no. 1, p. 185-208.
Br�nnimann P. (1940).- �ber die terti�ren Orbitoididen und die Miogypsiniden von Nordwest Marokko.- Schweizerische Pal�ontologische Abhandlungen, Basel, vol. 63, 113 p.
Bryan J.R. (1995).- Life history and development of Oligocene Larger Benthic Foraminifera: A test of the environmental control on heterochrony.- Tulane Studies in Geology and Paleontology, New Orleans - LA, vol. 27, p. 101-118. URL: https://journals.tulanE.edu/tsgp/article/view/667/585
Cahuzac B. & Poignant A. (1991).- Morphologie des esp�ces de Pararotalia et de Miogypsinoides (foraminiferida) dans l'Oligoc�ne d'Aquitaine m�ridionale.- G�obios, Villeurbanne, vol. p. 24, p. 69-78.
Cahuzac B. & Poignant A. (1997).- Essai de biozonation de l'Oligo-Mioc�ne dans les bassins europ�ens � l'aide des grands foraminif�res n�ritiques.- Bulletin de la Soci�t� G�ologique de France, Paris, vol. 168, no. 2, p. 155-169.
Cerutti P. (1973, unpublished).- Macroforaminiferi della sezione del Rio Civera nella Collina di Torino.- BSc thesis, University of Torino [Not seen: data from Ferrero et al., 1994].
Cole W.S. (1938).- Stratigraphy and micropaleontology of two deep wells in Florida.- Florida State Geological Survey, Geological Bulletin, Tallahassee - FL, no. 16, p. 1-73. URL: http://ufdc.ufl.edu/UF00000453/00001
Cole W.S. (1957).- Late Oligocene Larger Foraminifera from Barro Colorado Island, Panama Canal Zone (with a detailed analysis of American Miogypsinids and Heterosteginids).- Bulletins of American Paleontology, Ithaca - NY, vol. XXXVII, no. 163, p. 313-338. URL: https://www.biodiversitylibrary.org/item/40446
Cole W.S. (1964).- American mid-Tertiary miogypsinid foraminifera: Classification and zonation.- Contributions from the Cushman Foundation for Foraminiferal Research, Glen Allen - VA, vol. XV, no. 4, p. 138-161. URL: https://business.cushmanfoundation.org/ap/CloudFile/Download/PdKzdBgp
Cole W.S. (1967).- A review of American species of miogypsinids (Larger Foraminifera).- Contributions from the Cushman Foundation for Foraminiferal Research, Glen Allen - VA, vol. XVIII, no. 3, p. 99-117. URL: https://business.cushmanfoundation.org/ap/CloudFile/Download/LE1QnxGL
Cushman J.A. (1918).- The larger fossil Foraminifera of the Panama Canal Zone.- Smithsonian Institution, United States National Museum, Bulletin, Washington - DC, no. 103, p. 89-102. URL: https://www.biodiversitylibrary.org/page/32168416
Cushman J.A. (1919).- Fossil foraminifera from the West Indies. In: Vaughan T.W. (ed.), Contributions to the geology and paleontology of the West Indies.- Carnegie Institution of Washington, Publication, no. 291, p. 21-71. URL: https://www.biodiversitylibrary.org/item/69909
Delicati S. & Schiavinotto F. (1985).- Miogypsinidae from the Miocene of Monte Torretta (L'Aquila, Centrai Apennines): Taxonomy, phylogeny and biostratigraphic significance.- Geologica Romana, vol. XXIV, p. 101-146 [Not seen: data from Ferrero et al., 1994].
Dervieux E. (1894).- Osservazioni sopra le Tinoporinae e descrizione del nuovo genre Flabelliporus.- Atti dell'Accademia della Scienze, Torino, vol. 29, p. 57-61.
Drooger C.W. (1951).- Notes on some representatives of Miogypsinella.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LIV, p. 357-365. URL: https://archive.org/download/proceedings-knaw-series-b_1951_54/proceedings-knaw-series-b_1951_54.pdf
Drooger C.W. (1952).- Study of American Miogypsinidae.- Vonk & Co's Drukkeeij, Zeist, 80 p.
Drooger C.W. (1954a).- Miogypsina in northern Italy.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LVII, p. 227-249. URL: https://archive.org/download/proceedings-knaw-series-b_1954_57/proceedings-knaw-series-b_1954_57.pdf
Drooger C.W. (1954b).- Miogypsina in Northwestern Morocco.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LVII, p. 580-591. URL: https://archive.org/download/proceedings-knaw-series-b_1954_57/proceedings-knaw-series-b_1954_57.pdf
Drooger C.W. (1963).- Evolutionary trends in Miogypsinidae. In: Vlerk I.M. van der (ed.), Evolutionary trends in Foraminifera: A collection of papers dedicated to I.M. van der Vlerk on the occasion of his 70th birthday.- Elsevier, Amsterdam, p. 315-349.
Drooger C.W. (1993).- Radial Foraminifera, morphometrics and evolution.- Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afdeling Natuurkunde (Eerste Reeks), Amsterdam, deel 41, 242 p.
Drooger C.W. & Freudenthal T. (1964).- Associations of Miogypsina and Lepidocyclina at some European localities.- Eclogae Geologicae Helvetiae, Basel, vol. 57, p. 509-528. DOI: 10.5169/seals-163147
Drooger C.W. & Laagland H. (1986).- Larger foraminiferal zonation of the European Mediterranean Oligocene.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LXXXIX, p. 135-148.
Drooger C.W. & Raju D.S.N. (1973).- Protoconch diameter in the Miogypsinidae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LXXVI, p. 206-216.
Drooger C.W. & Raju D.S.N. (1978).- Early Miogypsinoides in Kutch, Western India (I - II). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Serie B), Amsterdam, vol. LXXXI, p. 186-219.
Drooger C.W., Kaasschieter J.P.H. & Key A.J. (1955).- The microfauna of the Aquitanian-Burdigalian of southwestern France.- Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afdeling Natuurkunde (Eerste Reeks), Amsterdam, deel 21 no. 2, p. 1-136 [Not seen: data from Ferrero et al., 1994].
Dunham R.J. (1962).- Classification of carbonate rocks according to depositional texture. In: Ham W.E. (ed.), Classification of Carbonate Rocks - A Symposium.- American Association of Petroleum Geologists, Memoir, Tulsa - OK, no. 1, p. 108-121.
Embry A.F. & Klovan J.E. (1971).- A Late Devonian reef tract on Northeastern Banks Island, NWT.- Bulletin of Canadian Petroleum Geology, Calgary, vol. 19, p. 730-781.
Eva A. & McFarlane N. (1985).- Tertiary to early Quaternary carbonate facies relationships in Jamaica.- Transactions of the Fourth Latin American Geological Congress Trinidad and Tobago, 7th-15th July, 1979, Arima.- Trinidad & Tobago Print & Packaging Ltd., Arima, p. 210-219.
Fauquembergue K., Mulder T., Reijmer J., Hanquiez V., Betzler C., Ducassou E., Recouvreur A., Principaud M., Borgomano J., Wilk S. & Poli E. (2024).- Sedimentology of modern Bahamian carbonate slopes: Summary and update.- Geochemistry, Geophysics, Geosystems, vol. 25, no. 6, 21 p. DOI: 10.1029/2023GC011077
Ferrero E. (1965).- Miogypsine della Formazione di Termofour� nella serie del Rio di Superga (Collina di Torino).- BSc thesis, University of Torino [Not seen: data from Ferrero et al., 1994].
Ferrero E. (1987, unpublished).- Miogypsinidi della serie oligo-miocenica della Collina di Torino (Italia Nord-Occidentale).- Bollettino della Societ� Paleontologia Italiana, Torino, vol. 26, nos. 1-2, p. 119-150. URL: https://www.paleoitalia.it/wp-content/uploads/2024/06/08_Mortara_1987_BSPI_261-2.pdf
Ferrero E., Maia F. & Tonon M. (1994).- Le Miogypsine del Monferrato: Aspetti morfologici e tassonomici.- Bollettino della Societ� Paleontologia Italiana, Torino, vol. 33, no. 3, p. 345-368. URL: https://www.paleoitalia.it/wp-content/uploads/2024/07/04_Ferrero-et-al_1994_BSPI_333.pdf
Frost S.H. & Langenheim R.L. (1974).- Tertiary larger foraminifera and scleractinian corals from Mexico. In: Cenozoic reef biofacies.- Northern Illinois University Press, DeKalb - IL, 388 p.
Gravell D.W. (1933).- Tertiary larger foraminifera of Venezuela.- Smithsonian Miscellaneous Collections, Washington - DC, vol. 89, no. 11, p. 1-44. URL: https://repository.si.edu/bitstreams/277135a2-74c2-4b11-97ef-a9349db33d9d/download
G�mbel C.W. von (1870).- Beitr�ge zur Foraminiferenfauna der nordalpinen, �lteren Eoc�ngebilde oder der Kressenberger Nummulitenschichten.-
Abhandlungen der Mathematisch-Physikalischen Klasse der K�niglich Bayerischen Akademie der
Wissenschaften, M�nchen, vol. 10, no. 2, p. 581-730. URL: https://mdz-nbn-resolving.de/urn:nbn:de:bvb:12-bsb10226029-2 Hanzawa S. (1940).- Micropalaeontological studies of drill cores from a deep well in Kita-Daito-Zima (North Borodino lsland). In: Jubilee Publication in the Commemoration of Professor H. Yabe, M.I.A. Sixtieth Birthday, Volume 2.- Sasake Publishing Co., Sendai, p.
755-802 [Not seen]. Hanzawa S. (1968).- [Larger foraminifera].- Asakura Shoten, Tokyo, 300
p. [Not seen, in Japanese]. Hodson H.K. (1926).- Foraminifera from Venezuela and Trinidad.- Bulletins of American Paleontology, Ithaca - NY, vol. XII, no. 47, p. 1-46. URL:
https://www.biodiversitylibrary.org/item/95757 Hose H.R. & Versey H.R. (1956).- Palaeontological and lithological divisions of the lower Tertiary limestones of Jamaica.- Colonial Geology and Mineral Resources, London, vol. 6, p. 19-39. Hottinger L., Halicz E. & Reiss Z. (1991).- The foraminiferal genera Pararotalia, Neorotalia, and Calcarina: Taxonomic revision.- Journal of Paleontology, Tulsa - OK, vol. 65, no. 1, p. 18-33. Koch R. (1926).- Miogypsina staufferi, nov. spec., from north-western Venezuela.- Eclogae Geologicae Helvetiae, Basel, vol. 19, no. 3, p. 751-753. DOI:
10.5169/seals-158435 Land L.S. (1991).- Some aspects of the late Cenozoic evolution of north Jamaica as revealed by strontium isotope stratigraphy.- Journal of the Geological Society of Jamaica, vol. 28, p. 45-48. Loeblich A.R. Jr & Tappan H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold Co., New York - NY, 730 p. Lokier S.W. & Al Junaibi M. (2016).- The petrographic description of carbonate facies:
Are we all speaking the same language?- Sedimentology, vol. 63, p. 1843-1885. DOI: 10.1111/sed.12293 Lopez-Gamundim C., Barnes B.B, Betzler C., Harris P.M., Oehlert A.M., Eberli G.P. & Purkis S.J. (2025).- The sediment budget of Great Bahama Bank-Earth's largest modern carbonate platform.- Geology, Boulder
- CO, vol. 53, no. 9, p. 748-752. Lunt P. & Luan X. (2022).- SE Asian Cenozoic Larger Foraminifera: Taxonomic questions, apparent radiation and abrupt extinctions.- Journal of Earth Science, vol. 33, no. 6, p. 1378-1399. Matsumaru K. (1990).- A new genus of the miogypsinid foraminifer from southwest Japan.- Transactions and Proceedings of the Paleontological Society of Japan (New series), Tokyo, vol. 158, p. 535-539. DOI:
10.14825/prpsj1951.1990.158_535 Matsumaru K. (1996).- Tertiary larger foraminifera (Foraminiferida) from the Ogasawara Islands, Japan.- Paleontological Society of Japan, Special Paper, Tokyo, no. 36, 239 p. McFarlane N. (compiler) (1977).- 1:250,000 Jamaica. Geology.- Mines and Geology Division, Ministry of Mining and Natural Resources, Kingston (Jamaica). Michelotti G. (1841).- Saggio storico dei Rizopodi caratteristici dei terreni sopracretacei.- Memorie della Societ� Italiana delle Scienze, Modena, t. XXII, p. 253-302. URL:
https://natuurtijdschriften.nl/pub/541700 Mitchell S.F. (2004).- Lithostratigraphy and palaeogeography of the White Limestone Group. In: Donovan S.K. (ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Leiden, vol. 3, vol. 3, nos. 1-2, p. 5-29. URL:
https://natuurtijdschriften.nl/pub/541700 Mitchell S.F. (2013).- Stratigraphy of the White Limestone of Jamaica.- Bulletin de la Soci�t� G�ologique de France, Paris, vol. 184, nos. 1-2, p. 111-118. Mitchell S.F., Robinson E., �zcan E., Jiang M.M. & Robinson N. (2022).- A larger benthic foraminiferal zonation for the Eocene of the Caribbean and central American region.- Carnets Geol., Madrid, vol. 22, no. 11, p. 409-565.
DOI: 10.2110/carnets.2022.2211 Mitchell S.F., Robinson E., �zcan E., Jiang M.M. & Robinson N. (2024).- A larger benthic foraminiferal zonation for the Cenozoic of the Americas.- Carnets Geol., Madrid, vol. 24, no. 10, p. 163-172. DOI:
10.2110/carnets.2024.2410 Mulder E.F.J. de (1975).- Microfauna and sedimentary-tectonic history of the Oligo-Miocene of the Jonian Islands and Western Epirus (Greece).- Utrecht Micropaleontological Bulletins, vol. 13, 140 p. URL:
https://dspace.library.uu.nl/items/fec73e90-1378-42d0-a4a7-99a82953f9f7 Nigam R. & Rao A.S. (1987).- Proloculus size variation in recent benthic Foraminifera: Implications for paleoclimatic studies.- Estuarine, Coastal and Shelf Science, vol. 24, no. 5, p. 649-655. Novandaru N. Maryunani K.A., Kapid R., Aswan A. & Khorniawan W.B. (2025).- Phylogenetic evolution of Neorotalia mecatepecensis to Miogypsinidae from Rajamandala Formation, West Java, Indonesia.- Palaeoworld, vol. 34, no. 5, article 200961, 12 p. Nuttall W.L.F. (1928).- Notes on the Tertiary Foraminifera of Southern Mexico.- Journal of Paleontology, Tulsa - OK, vol. 2, no. 4, p. 372-376. Nuttall W.L.F. (1933).- Two species of Miogypsina from the Oligocene of Mexico.- Journal of Paleontology, Tulsa - OK, vol. 7, p. 175-177. Orbigny A.D. d' (1826).- Tableau m�thodique de la classe des C�phalopodes.- Annales des Sciences Naturelles, Paris, vol. 7, p. 96-169, 245-314. �zcan E., Less G., B�ldi-Beke M. & Koll�ny K. (2010).- Oligocene hyaline larger foraminifera from Kelereşdere Section (Muş, Eastern Turkey).- Micropaleontology, Flushing - NY, vol. 56, no. 5, p. 465-493. Raffi I., Wade B.S. & P�like H. with contributions by Beu A.G., Cooper R., Crundwell M.P., Krijgsman W., Moore T., Raine I., Sardella R. & Vernyhorova Y.V. (2020).- The Neogene Period. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale.- Elsevier, Amsterdam, p. 1141-1215. Raju D.S.N. (1974).- Study of Indian Miogypsinidae.- Utrecht Micropaleontological Bulletins, vol. 9, 148 p. URL:
http://hdl.handle.net/1874/205813 Reiss Z. (1963).- Reclassification of perforate foraminifera.- Bulletin of the Geological Survey of Israel, Jerusalem, vol. 35, p. 1-111. Robinson E. (1969,
unpublished).- Studies in the Tertiary stratigraphy of Eastern Jamaica.- PhD thesis, University of London, 289 p. Robinson E. (2004).- Zoning the White Limestone Group of Jamaica using larger foraminiferal genera: A review and proposal. In: Donovan S.K. (ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Leiden, vol. 3, nos. 1-2, p. 39-75. URL:
https://natuurtijdschriften.nl/pub/541702 Robinson E. & Wright R.M. (1993).- Jamaican Paleogene larger foraminifera. In: Wright R.M. & Robinson E. (eds.), Biostratigraphy of Jamaica.- Geological Society of America, Memoir, Boulder - CO, no. 182, p. 283-345. Robinson E. & Mitchell S.F. (1999).- Middle Eocene to Oligocene stratigraphy and palaeogeography in Jamaica: A window on the Nicaragua Rise.- Contributions to Geology UWI Mona, Kingston (Jamaica), no. 4, 47 p. URL:
http://www.sfmgeology.com/pdfs/MiddleEocenetoOligoceneStratigraphy.pdf Robinson E., Paytan A. & Chien C.-T. (2017).- Strontium isotope dates for the Oligocene Antigua Formation, Antigua, W.I.- Caribbean Journal of Earth Science, Kingston (Jamaica),vol. 50, p. 11-18. URL:
http://caribjes.com/CJESpdf/CJES50-3-Robinson-Antigua.pdf Robinson E., Paytan A., Chien C.-T. & Broach K. (2018).- Dating the White Limestone of Jamaica using Sr isotope stratigraphy: A progress report.- Caribbean Journal of Earth Science, Kingston (Jamaica), vol. 49, p. 11-21. URL:
https://caribjes.com/CJESpdf/CJES49-2-RobinsonSrIsotopes.pdf Rutten L.M.R. (1911).- On Orbitoides in the neighboorhood of the Balik Papan Bay, east-coast of Borneo.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, vol. XIII, 2nd part, p. 1122-1139. URL:
https://dwc.knaw.nl/DL/publications/PU00013345.pdf Sacco F. (1893).- Sur quelques Tinoporinae du Mioc�ne de Turin.- Bulletin de la Soci�t� Belge de G�ologie, de Pal�ontologie et d'Hydrologie, Bruxelles, t. VII, p. 204-207. URL:
https://archive.org/details/sacc_surq_1893 Salmer�n P. (1972).- Mutaci�n entre los g�neros Pararotalia y Miogypsinoides.- Revista del Instituto Mexicano del Petr�leo, Ciudad de M�xico, vol. 4, no. 3, p. 5-27. Schiavinotto F. (1979).- Miogypsina e Lepidocyclina del Miocene di Monte La Serra (L'Aquila, Appennino Centrale).- Geologica Romana, vol. 18, p. 253-293. Schiavinotto F. (1985).- Different evolutionary stages in the Miogypsinidae from Sardinia.- Bollettino della Societ� Paleontologia Italiana, Modena, vol. 23, no. 2, p. 381-393. URL:
https://www.paleoitalia.it/wp-content/uploads/2023/06/017_Schiavinotto.pdf Schlumberger C. (1900).- Note sur le genre Miogypsina.- Bulletin de la Soci�t� g�ologique de France (3e s�rie), Paris, t. XXVIII, no. 3, p. 327-333. URL:
https://www.biodiversitylibrary.org/page/30547825 Sch�ttenhelm R.T.E. (1976).- History and modes of Miocene carbonate deposition in the interior of the Piedmont Basin, NW Italy.- Utrecht micropaleontological Bulletins, no. 14, 197 p. URL:
http://hdl.handle.net/1874/205818 Silvestri A. (1907).- Probabile origine d'alcune Orbitoidine.- Rivista Italiana di Paleontologia, Perugia, vol. XIII, p. 79-81. URL:
https://archive.org/Sirel/rivistaitalianad13mila#page/79/ Sirel E. & Gedik F. (2011).- Postmiogypsinella, a new Miogypsinidae (Foraminifera) from the Late Oligocene in Malatya Basin, Turkey.- Revue de Pal�obiologie, Gen�ve, vol. 30, no. 2, p. 591-603. Soediono H. (1969).- Planktonic Foraminifera from the Velez Rubio Region, SE Spain. Part 1: The Ciudad Granada Formation (Oligo-Miocene).- Revista Espa�ola de Micropaleontolog�a, Madrid, vol. 1, no. 3, p. 335-353. Stemann T.A. (2004).- Reef corals of the White Limestone Group of Jamaica.-
Cainozoic Research, vol. 3, nos. 1-2, p. 83-107. URL: https://natuurtijdschriften.nl/pub/541704 Tan Sin Hok (1936).- Zur Kenntniss der Miogypsiniden.- De Ingenieur in Nederlandsch-Indi�, Bandung, III (3e Jaargang), no. 3, p. 45-61. URL:
https://www.stichtingblauwelijn.nl/assets/files/1936-03.pdf; vol. 3, no. 5, p. 84-98. URL:
https://www.stichtingblauwelijn.nl/assets/files/1936-05.pdf; vol. 3, no. 7, p. 109-123. URL:
https://www.stichtingblauwelijn.nl/assets/files/1936-07.pdf Tan Sin Hok (1937).- Weitere Untersuchungen �ber die Miogypsiniden I.- De Ingenieur in Nederlandsch-Indi�, Bandung, IV (4e Jaargang), no. 3, p. 35-45. URL:
https://www.stichtingblauwelijn.nl/assets/files/1937-03.pdf; no. 6, p. 87-111. URL:
https://www.stichtingblauwelijn.nl/assets/files/1937-06.pdf Vaughan T.W. (1928a).- Species of large arenaceous and orbitoidal Foraminifera from the Tertiary deposits of Jamaica.- Journal of Paleontology, Tulsa - OK, vol. 1, no. 4, p. 277-298. Vaughan T.W. (1928b).- Subfamily Miogypsininae Vaughan. In: Cushman J.A. (ed.), Foraminifera their classification and economic use.- Special Publications Cushman Laboratory for Foraminiferal Research, Sharon - MA, no. 1, p. 354-355 (Pls. 56-57, 59). URL:
https://archive.org/embed/foraminiferathei00cush Vlerk I.M. van der (1924).- Miogypsina Dehaartii, nov. spec. de Larat (Moluques).- Eclogae Geologicae Helvetiae, Basel, vol. XVIII, Heft 3, p. 429-432. DOI:
10.5169/seals-158257 Wildenborg A.F.B. (1991).- Evolutionary aspects of the Miogypsinids in the Oligo-Miocene carbonates near Mineo (Sicily).- Utrecht micropaleontological Bulletins, no. 41, 139 p. URL:
https://dspace.library.uu.nl/handle/1874/205900 Yu Zhoufei, Li Yanli & Li Tiegang (2016).- Mean proloculus size as a salinity index in benthic Foraminifera Ammonia aomoriensis: Based on culture and seasonal studies.- Journal of the Palaeontological Society of India, Lucknow, vol. 61, no. 2, p. 215-223. DOI:
10.1177/0971102320160204 Zans V.A., Chubb L.J., Versey H.R., Williams J.B., Robinson E. & Cooke D.L. (1963).- Synopsis of the geology of Jamaica. An explanation of the 1958 provisional geological map of Jamaica.- Geological Survey Department, Bulletin, Kingston (Jamaica), no. 4, 72
p. [Dated on front cover and title page 1962, Printers imprint at bottom of front page is 1963]. Data on Jamaica miogypsinids used in this work.
Appendix
Sample
Slab
No
Formation
Location
Species
X
γ
Pw
R1118
...
T/S
Chester Fm
Sherwood Content
complanatus
20
-269
108
R1118
...
T/S
Chester Fm
Sherwood Content
complanatus
18
-297
118
R1118
Slab 1
3
Chester Fm
Sherwood Content
complanatus
15
-213
110
R1118
Slab 1
4
Chester Fm
Sherwood Content
complanatus
21
-380
115
R1118
Slab 1
5
Chester Fm
Sherwood Content
complanatus
18
-240
113
R1118
Slab 1
6
Chester Fm
Sherwood Content
complanatus
20
-370
96
R1118
Slab 1
7
Chester Fm
Sherwood Content
complanatus
17
-252
115
R1118
Slab 1
8
Chester Fm
Sherwood Content
complanatus
20
-310
92
R1118
Slab 1
9
Chester Fm
Sherwood Content
complanatus
17
-201
106
R1118
Slab 1
10
Chester Fm
Sherwood Content
complanatus
17
-285
125
R1118
Slab 1
11
Chester Fm
Sherwood Content
complanatus
16
-275
116
R1118
Slab 1
12
Chester Fm
Sherwood Content
complanatus
17
-273
R1118
Slab 1
13
Chester Fm
Sherwood Content
complanatus
19
138
R1118
Slab 2a
14
Chester Fm
Sherwood Content
complanatus
20
-304
137
R1118
Slab 2a
15
Chester Fm
Sherwood Content
complanatus
18
-274
105
R1118
Slab 2b
16
Chester Fm
Sherwood Content
complanatus
16
-231
107
R1118
Slab 2b
17
Chester Fm
Sherwood Content
complanatus
21
-375
115
R1118
Slab 3a
18
Chester Fm
Sherwood Content
complanatus
17
-281
88
R1118
Slab 3a
19
Chester Fm
Sherwood Content
complanatus
17
-285
121
R1118
Slab 3a
20
Chester Fm
Sherwood Content
complanatus
16
-320
120
R1118
N/A
21
Chester Fm
Sherwood Content
complanatus
17
-261
R1118
N/A
22
Chester Fm
Sherwood Content
complanatus
15
-185
129
R1118
N/A
23
Chester Fm
Sherwood Content
complanatus
20
-336
107
R1118
N/A
24
Chester Fm
Sherwood Content
complanatus
20
-353
109
R1118
N/A
25
Chester Fm
Sherwood Content
complanatus
18
-294
124
R1118
N/A
26
Chester Fm
Sherwood Content
complanatus
18
-348
118
R1118
N/A
27
Chester Fm
Sherwood Content
complanatus
18
-317
113
R1118
N/A
28
Chester Fm
Sherwood Content
complanatus
15
-260
111
R1118
N/A
29
Chester Fm
Sherwood Content
complanatus
20
-272
R1118
N/A
30
Chester Fm
Sherwood Content
complanatus
19
-327
94
WL119
N/A
1
Basal Newport Fm
Highway 2000
tani
9
-31
WL119
N/A
2
Basal Newport Fm
Highway 2000
tani
8
-32
138
WL119
N/A
3
Basal Newport Fm
Highway 2000
tani
10
-18
128
WL119
N/A
4
Basal Newport Fm
Highway 2000
tani
7
-39
123
WL1727
N/A
3
Chester Fm
St Mary
complanatus
6
-8
114
WL1727
N/A
4
Chester Fm
St Mary
complanatus
6
-12
128
WL1727
N/A
5
Chester Fm
St Mary
complanatus
6
-21
140
WL1727
N/A
6
Chester Fm
St Mary
complanatus
8
-85
123
WL1727
N/A
9
Chester Fm
St Mary
complanatus
8
-28
135
WL1727
N/A
10
Chester Fm
St Mary
complanatus
8
0
130
WL1727
N/A
12
Chester Fm
St Mary
complanatus
7
-53
114
WL1727
N/A
13
Chester Fm
St Mary
complanatus
9
-9
118
WL1727
N/A
14
Chester Fm
St Mary
complanatus
7
-40
117
WL1727
N/A
16
Chester Fm
St Mary
complanatus
8
-53
134
WL1727
N/A
17
Chester Fm
St Mary
complanatus
7
7
130
WL1727
N/A
18
Chester Fm
St Mary
complanatus
6
0
118
WL1727
N/A
19
Chester Fm
St Mary
complanatus
8
-31
125
WL1727
N/A
21
Chester Fm
St Mary
complanatus
8
-21
119
WL3669
Slab3
1
Basal Newport Fm
Ulster Spring
tani
8
-61
145
WL3669
Slab3
2
Basal Newport Fm
Ulster Spring
tani
9
-71
139
WL3669
Slab3
3
Basal Newport Fm
Ulster Spring
tani
8
-60
146
WL3669
Slab 4
4
Basal Newport Fm
Ulster Spring
tani
10
-97
106
WL3669
Slab 1
5
Basal Newport Fm
Ulster Spring
tani
9
-193
100
WL3669
Slab 1
6
Basal Newport Fm
Ulster Spring
tani
9
-52
119
WL3669
Slab 2a
7
Basal Newport Fm
Ulster Spring
tani
7
-51
112
WL3669
Slab 2a
8
Basal Newport Fm
Ulster Spring
tani
7
-38
110
WL3669
Slab 2a
9
Basal Newport Fm
Ulster Spring
tani
7
-58
WL3669
Slab 2a
10
Basal Newport Fm
Ulster Spring
tani
8
-76
107
WL3669
Slab 2a
11
Basal Newport Fm
Ulster Spring
tani
10
-48
129
WL3669
Slab 2a
12
Basal Newport Fm
Ulster Spring
tani
8
-42
127
WL3669
Slab 2a
12A
Basal Newport Fm
Ulster Spring
tani
10
-51
119
WL3669
Slab 2c
13
Basal Newport Fm
Ulster Spring
tani
8
-35
WL3669
Slab 2b
14
Basal Newport Fm
Ulster Spring
tani
8
-48
142
WL3669
Slab 2b
15
Basal Newport Fm
Ulster Spring
tani
7
-44
131
WL3669
Slab 2b
16
Basal Newport Fm
Ulster Spring
tani
7
-53
113
WL3669
Slab 3A
16
Basal Newport Fm
Ulster Spring
tani
8
-35
121
WL3676
N/A
1
Moneague Fm
Ulster Spring
basraensis
13
-216
118
WL3676
N/A
2
Moneague Fm
Ulster Spring
basraensis
13
-212
91
WL3676
N/A
3
Moneague Fm
Ulster Spring
basraensis
17
-212
77
WL3676
N/A
4
Moneague Fm
Ulster Spring
basraensis
14
-218
120
WL3807
N/A
1
Moneague Fm
Ulster Spring
complanatus
18
-299
59
WL3807
N/A
2
Moneague Fm
Ulster Spring
complanatus
17
-221
69
WL3807
N/A
3
Moneague Fm
Ulster Spring
complanatus
19
-274
97
WL3807
N/A
4
Moneague Fm
Ulster Spring
complanatus
21
-328
85
WL3807
N/A
5
Moneague Fm
Ulster Spring
complanatus
17
-279
82
WL3807
N/A
6
Moneague Fm
Ulster Spring
complanatus
16
-259
104
WL3807
N/A
7
Moneague Fm
Ulster Spring
complanatus
18
-301
138
WL3807
N/A
8
Moneague Fm
Ulster Spring
complanatus
21
-344
133
WL3807
N/A
9
Moneague Fm
Ulster Spring
complanatus
19
-292
92
WL3807
N/A
10
Moneague Fm
Ulster Spring
complanatus
19
-345
69
WL3809
Slab 1 bottom
1
Moneague Fm
Ulster Spring
thalmanni
16
-265
100
WL3809
Slab 1 bottom
2
Moneague Fm
Ulster Spring
thalmanni
17
-325
133
WL3809
Slab 1 bottom
4a
Moneague Fm
Ulster Spring
thalmanni
16
-232
WL3809
Salb 1 top
5
Moneague Fm
Ulster Spring
thalmanni
13
-169
125
WL3809
Salb 1 top
6
Moneague Fm
Ulster Spring
thalmanni
14
-226
139
WL3809
Salb 1 top
7
Moneague Fm
Ulster Spring
thalmanni
17
-246
WL3809
Salb 1 top
8
Moneague Fm
Ulster Spring
thalmanni
16
-210
97
WL3809
Salb 1 top
9
Moneague Fm
Ulster Spring
thalmanni
14
-237
97
WL3809
Slab 2
10
Moneague Fm
Ulster Spring
thalmanni
13
-149
110
WL3809
Slab 2
11
Moneague Fm
Ulster Spring
thalmanni
19
-420
WL3809
Slab 2
12
Moneague Fm
Ulster Spring
thalmanni
14
-237
100
WL3809
Slab 3
13
Moneague Fm
Ulster Spring
thalmanni
15
-159
94
WL3809
Slab 3
14
Moneague Fm
Ulster Spring
thalmanni
15
-233
97
WL3809
Slab 3
15
Moneague Fm
Ulster Spring
thalmanni
16
-267
116
WL3809
Slab 3
16
Moneague Fm
Ulster Spring
thalmanni
14
-257
120
WL3809
Slab 3
17
Moneague Fm
Ulster Spring
thalmanni
17
-243
95
WL3809
Slab 4
18
Moneague Fm
Ulster Spring
thalmanni
17
-262
111
WL3809
Slab 4
19
Moneague Fm
Ulster Spring
thalmanni
17
-242
WL3809
Slab 4
20
Moneague Fm
Ulster Spring
thalmanni
16
-239
86
WL3809
Slab 4
21
Moneague Fm
Ulster Spring
thalmanni
14
-199
115
WL3809
Slab 4
21a
Moneague Fm
Ulster Spring
thalmanni
14
-157
136
WL3810
Slab 1
1
Moneague Fm
Ulster Spring
basraensis
11
-141
113
WL3810
Slab 1
2
Moneague Fm
Ulster Spring
basraensis
12
-137
117
WL3810
Slab 1
3
Moneague Fm
Ulster Spring
basraensis
13
-209
108
WL3810
Slab 1
4
Moneague Fm
Ulster Spring
basraensis
13
-211
119
WL3810
Slab 1
5
Moneague Fm
Ulster Spring
basraensis
12
-169
120
WL3810
Slab 1
6
Moneague Fm
Ulster Spring
basraensis
12
-224
98
WL3810
Slab 1
7
Moneague Fm
Ulster Spring
basraensis
13
-162
133
WL3810
Slab 1
8
Moneague Fm
Ulster Spring
basraensis
11
-214
WL3810
Slab 1
12
Moneague Fm
Ulster Spring
basraensis
12
-186
143
WL3810
Slab 1
13
Moneague Fm
Ulster Spring
basraensis
13
-145
96
WL3810
Slab 2a
14
Moneague Fm
Ulster Spring
basraensis
12
-186
89
WL3810
Slab 2a
15
Moneague Fm
Ulster Spring
basraensis
10
-122
147
WL3810
Slab 2b
16
Moneague Fm
Ulster Spring
basraensis
14
-195
134
WL3810
Slab 2b
17
Moneague Fm
Ulster Spring
basraensis
15
-180
114
WL3810
Slab 3a
18
Moneague Fm
Ulster Spring
basraensis
13
-111
131
WL3810
Slab 3a
19
Moneague Fm
Ulster Spring
basraensis
13
-163
120
WL3810
Slab 3a
20
Moneague Fm
Ulster Spring
basraensis
12
-195
127
WL3810
Slab 3a
21
Moneague Fm
Ulster Spring
basraensis
14
-179
WL3810
Slab 3a
22
Moneague Fm
Ulster Spring
basraensis
14
-241
100
WL3810
Slab 3b
23
Moneague Fm
Ulster Spring
basraensis
13
-257
113
WL3810
Slab 3b
24
Moneague Fm
Ulster Spring
basraensis
12
-178
89
WL3810
Slab 3b
25
Moneague Fm
Ulster Spring
basraensis
11
-180
92
WL3810
Slab 3b
26
Moneague Fm
Ulster Spring
basraensis
16
-256
122
WL3810
Slab 3b
27
Moneague Fm
Ulster Spring
basraensis
12
-150
124
WL3810
Slab 3b
28
Moneague Fm
Ulster Spring
basraensis
14
-279
WL3810
Slab 3b
29
Moneague Fm
Ulster Spring
basraensis
15
-188
133
WL4791
Slab4
...
Moneague Fm
St Ann
complanatus
18
-150
WL4791
Slab4
...
Moneague Fm
St Ann
complanatus
16
-199
106
WL4791
Slab4
...
Moneague Fm
St Ann
complanatus
19
-281
90
WL4791
Slab4
...
Moneague Fm
St Ann
complanatus
18
-328
112
WL4791
N/A
...
Moneague Fm
St Ann
complanatus
23
-375
95
WL4791
N/A
...
Moneague Fm
St Ann
complanatus
22
-395
87
WL4791
N/A
...
Moneague Fm
St Ann
complanatus
20
-356
102
WL497
Slab 1
1
Moneague Fm
Prickly Pole
basraensis
14
-213
114
WL497
Slab 1
2
Moneague Fm
Prickly Pole
basraensis
14
-161
153
WL497
Slab 1
3
Moneague Fm
Prickly Pole
basraensis
12
-165
108
WL497
Slab 1
4
Moneague Fm
Prickly Pole
basraensis
13
-150
118
WL497
Slab 2
b
Moneague Fm
Prickly Pole
basraensis
14
-251
WL497
Slab 2
5
Moneague Fm
Prickly Pole
basraensis
15
-232
88
WL497
Slab 3
1
Moneague Fm
Prickly Pole
basraensis
12
-159
113
WL497
Slab 4
1
Moneague Fm
Prickly Pole
basraensis
14
-250
104
WL497
Slab 5
1
Moneague Fm
Prickly Pole
basraensis
12
-133
116
WL497
Slab 6
1
Moneague Fm
Prickly Pole
basraensis
15
-232
92
WL497
Slab 6
2
Moneague Fm
Prickly Pole
basraensis
14
-169
108
WL5008
N/A
9
Chester Fm
Hope Bay
tani
5
-12
138
WL5008
N/A
10
Chester Fm
Hope Bay
tani
6
-39
201
WL5008
N/A
20
Chester Fm
Hope Bay
tani
6
9
158
WL5008
N/A
22
Chester Fm
Hope Bay
tani
8
-53
148
WL5008
N/A
23
Chester Fm
Hope Bay
tani
8
-37
154
WL5008
N/A
33
Chester Fm
Hope Bay
tani
7
-2
177
WL5008
N/A
34
Chester Fm
Hope Bay
tani
6
14
164
WL5008
N/A
35
Chester Fm
Hope Bay
tani
6
14
131
WL5008
N/A
36
Chester Fm
Hope Bay
tani
5
15
138
WL531
Slab 1
1
Moneague Fm
Prickly Pole
complanatus
19
-335
84
WL531
Slab 1
1
Moneague Fm
Prickly Pole
complanatus
17
-288
81
WL5548
Slab 1
1
Newport Fm
Maggotty
tani
8
-16
130
WL5548
Slab 1
2
Newport Fm
Maggotty
tani
10
-124
110
WL5548
Slab 1
3
Newport Fm
Maggotty
tani
6
-12
132
WL5548
Slab 1
4
Newport Fm
Maggotty
tani
8
-24
147
WL5548
Slab 1
5
Newport Fm
Maggotty
tani
6
-24
159
WL5548
Slab 1
6
Newport Fm
Maggotty
tani
7
-21
134
WL5548
Slab 1
7
Newport Fm
Maggotty
tani
7
-72
95
WL5548
Slab 1
8
Newport Fm
Maggotty
tani
8
-32
132
WL5548
Slab 1
9
Newport Fm
Maggotty
tani
10
-24
100
WL5548
Slab 1
10
Newport Fm
Maggotty
tani
7
-35
138
WL5548
Slab 1
11
Newport Fm
Maggotty
tani
8
-34
130
WL5548
Slab 2
1
Newport Fm
Maggotty
tani
7
6
131
WL5548
Slab 2
2
Newport Fm
Maggotty
tani
7
-24
145
WL5548
Slab 2
3
Newport Fm
Maggotty
tani
8
-28
126
WL5548
Slab 2
4
Newport Fm
Maggotty
tani
9
-32
129
WL5548
Slab 2
5
Newport Fm
Maggotty
tani
9
-38
119
WL5548
Slab 3
1
Newport Fm
Maggotty
tani
8
-43
162
WL5548
Slab 4
1
Newport Fm
Maggotty
tani
7
-18
126
WL5548
Slab 4
2
Newport Fm
Maggotty
tani
8
-84
107
WL5772
N/A
1
Chester Fm
Rio Bueno
complanatus
18
-272
119
WL5772
N/A
1
Chester Fm
Rio Bueno
complanatus
19
-344
87
WL5772
N/A
1
Chester Fm
Rio Bueno
complanatus
20
-346
93