◄ Carnets Geol. 26 (10) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Microfossils]
[5. Sedimentology and petrology]
[6. Systematic palaeontology]
[7. Discussion]
[8. Conclusions]
[Bibliographic references] and ...
[Appendix]
Czech Geological Survey, Klárov 3, 11821 Prague 1 (Czech Republic)
Institute of Geology and Palaeontology, Faculty of Science, Charles University, Albertov 6, 12843 Prague 2 (Czech Republic)
Czech Geological Survey, Klárov 3, 11821 Prague 1 (Czech Republic)
Institute of Geology and Palaeontology, Faculty of Science, Charles University, Albertov 6, 12843 Prague 2 (Czech Republic)
Published online in final form (pdf) on May 31, 2026
DOI 10.2110/carnets.2026.2610
![]()
[Editor: Bruno
R.C. Granier; language editor: Zev Brook]
![]()
The Biskoupky locality represents a tectonically delimited relict of Miaolingian sediments within Furongian volcanic rocks of the Křivoklát-Rokycany Volcanic Complex, serving as the southernmost outcrop of the Miaolingian Buchava Formation of the Skryje-Týřovice Basin. Over a century of research focused on this outcrop has produced a well-documented assemblage of comparatively rare remains of skeletal fauna, including species so far restricted to this locality; however, a microfossil analysis has until now never been conducted. Herein, we present the results of a palynological analysis of material from samples of fine-grained siltstone to shale from the Biskoupky locality. The samples yielded moderately well preserved organic-walled microfossils, particularly the acritarch genera Cristallinium, Eliasum, Heliosphaeridium, Retisphaeridium, Solisphaeridium, Synsphaeridium, and multiple species of Timofeevia. For the first time in the Skryje-Týřovice Basin, the presence of Adara? sp. is documented. The presence of Timofeevia here and in previously recorded acritarch assemblages from the Skryje-Týřovice Basin suggests a later-than-expected deposition of the Buchava Formation.
• acritarchs;
• Drumian;
• Miaolingian;
• biostratigraphy;
• Skryje-Týřovice Basin;
• Czech Republic
Kovář V., Vorel T. & Fatka O. (2026).- First palynological dating of Miaolingian (Cambrian) strata at the Biskoupky locality (Skryje-Týřovice Basin, Teplá-Barrandian unit).- Carnets Geol., Madrid, vol. 26, no. 10, p. 211-225. DOI: 10.2110/carnets.2026.2610
Premičre datation palynologique des strates du Miaolingien (Cambrien) de la localité de Biskoupky (Bassin de Skryje-Týřovice, unité de Teplá-Barrandien).- La localité de Biskoupky représente un vestige tectoniquement délimité de sédiments miaolingien au sein des volcanites furongiennes du Complexe Volcanique de Křivoklát-Rokycany. Elle constitue l'affleurement le plus méridional de la Formation de Buchava (Miaolingien) dans le Bassin de Skryje-Týřovice. Plus d'un sičcle de recherches menées sur cet affleurement ont permis de documenter un assemblage bien conservé de restes relativement rares de faune ŕ squelette, incluant des espčces exclusivement connues de cette localité ; cependant, une analyse des microfossiles n'avait jusqu'ŕ présent jamais été réalisée. Nous présentons ici les résultats d'une analyse palynologique conduite sur des échantillons de siltite ŕ grain fin et de schiste de la localité de Biskoupky. Les échantillons ont livré des microfossiles ŕ paroi organique modérément bien préservés, notamment des acritarches des genres Cristallinium, Eliasum, Heliosphaeridium, Retisphaeridium, Solisphaeridium et Synsphaeridium, ainsi que plusieurs espčces de Timofeevia. La présence d'Adara alea est documentée pour la premičre fois dans le Bassin de Skryje-Týřovice. La présence de Timofeevia, ici et dans les associations d'acritarches précédemment étudiées dans le Bassin de Skryje-Týřovice, suggčre un dépôt plus tardif que prévu pour la Formation de Buchava.
• acritarches ;
• Drumien ;
• Miaolingien ;
• biostratigraphie ;
• Bassin de Skryje-Týřovice ;
• République tchčque
In the Teplá-Barrandian unit (TBU), two major areas with
paleontologically dated Cambrian rocks have been known for more than two
centuries, the Příbram-Jince and the Skryje-Týřovice basins (e.g.,
Havlíček, 1971; Geyer et al.,
2008). In the
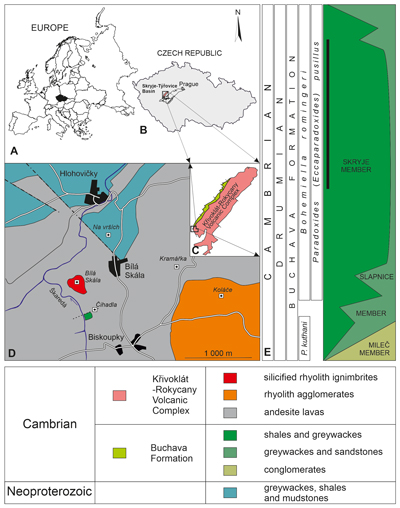
Skryje-Týřovice Basin, the Cambrian volcano-sedimentary sequence (Fig.
1.B-C ![]() ) rests unconformably on the slightly metamorphosed Neoproterozoic
rocks of the Cadomian consolidated basement. Sediments of the Miaolingian
Buchava Formation (Fatka et al., 2011b) are capped by Furongian
to Early Ordovician tholeiitic and calc-alkaline subaerial basalts, andesites
and rhyolites of the Křivoklát-Rokycany Volcanic Complex (Havlíček,
1971; Drost et al., 2004; Geyer et al.,
2008).
) rests unconformably on the slightly metamorphosed Neoproterozoic
rocks of the Cadomian consolidated basement. Sediments of the Miaolingian
Buchava Formation (Fatka et al., 2011b) are capped by Furongian
to Early Ordovician tholeiitic and calc-alkaline subaerial basalts, andesites
and rhyolites of the Křivoklát-Rokycany Volcanic Complex (Havlíček,
1971; Drost et al., 2004; Geyer et al.,
2008).
The Buchava Formation is represented by a sequence up to 200 metres thick, consisting of
conglomerates, sandstones, greywackes, and shales
cropping out in the northwestern part of the lower Paleozoic exposures of the
Barrandian area (Havlíček, 1971; Geyer et al.,
2008; Hajná et al., 2017). Cambrian sediments have been
recognized in a belt approximately 20 km long and up to 2 km wide. The
southernmost occurrence of skeletal fauna in the Buchava Formation has been
reported from a small outcrop near the village of Biskoupky (Fig.
1.C ![]() ).
).
The aim of this contribution is to describe and stratigraphically evaluate the recently discovered moderately well preserved organic-walled microfossils isolated from siltstones to shales of the Buchava Formation at the Biskoupky locality and to put the acritarch record of the Skryje-Týřovice Basin into a regional context.
In the mid-19th century, marine Cambrian sediments of the Skryje-Týřovice
Basin were classified as the "Ginetzer Schichten" (= Jince layers) by Lipold
and Krejčí (1860), based on the presence of the so-called
"faune
primordiale" of Barrande (1852), traditionally assigned to the étage
C of Barrande. Since then, numerous stratigraphic schemes have been proposed
for the strata. Most recently, the name Buchava Formation has been applied to
include all Cambrian sediments in the Skryje-Týřovice Basin by Fatka
et al. (2011b). These authors recognised three distinct units within the
Buchava Formation: the Skryje, Slapnice and Mileč members (Fig.
1.D ![]() ).
).
The fossil record of the Buchava Formation has been intensively studied for nearly 180 years (e.g., Barrande, 1846, 1852; Pompeckj, 1896; Šnajdr, 1958; Bruthansová et al., 2007). Locally common fossils have been collected from dozens of natural outcrops and excavations, both large and small, in the 19th and 20th centuries (for discussion, see Jahn, 1896; Šnajdr, 1958; Chlupáč, 1999; Fatka, 2004; Fatka et al., 2011b; Fatka & Herynk, 2016). Numerous articulated and disarticulated remains of trilobites and agnostids; stylophoran, cinctan, eocrinoid, and ctenocystid echinoderms; hyoliths; brachiopods; and rare bivalved arthropods, wiwaxiids, helcionelloid molluscs, pterobranchs, foraminifera, and ichnofossils have been documented. These fossils have been studied for nearly two hundred years (for a summary, see Fatka, 1990, 2004; Mikuláš, 2000; Maletz et al., 2005; Fatka et al., 2011a; Mikuláš & Fatka, 2017). In comparison, organic-walled microfossils have been reported only sporadically (Konzalová, 1974; Vavrdová, 1976; Chlupáč et al., 1998).
|
Figure 1:
Map showing the location of the outcrop near the village
of Biskoupky. A - Position of the Czech Republic in Europe. B - Czech Republic
with the Palaeozoic of the Teplá-Barrandian unit and the Skryje-Týřovice
area. C - Geological map showing the distribution of the middle Cambrian
Buchava Formation and the location of the Biskoupky locality. D - A simplified
geological map showing the position of the Biskoupky locality. E -
Stratigraphy of the Skryje-Týřovice Basin with a black bar representing
the supposed range of Mikaparia mutica. Geology modified from Vorel
et al. (2018); stratigraphy after Vokáč
(2002) and Fatka
et al. (2011b). |
Šnajdr
(1958) assigned the fossiliferous shales and greywackes to the Paradoxides (Eccaparadoxides)
pusillus Trilobite Biozone (Fig. 1.E ![]() ), drawing a direct comparison
between these strata and the lower portions of the Jince Formation (Příbram-Jince
Basin). Based on the stratigraphically restricted occurrence of brachiopods, Havlíček
(1971) defined the older Pompeckium kuthani and the younger Bohemiella
romingeri biozones (Fig. 1.E
), drawing a direct comparison
between these strata and the lower portions of the Jince Formation (Příbram-Jince
Basin). Based on the stratigraphically restricted occurrence of brachiopods, Havlíček
(1971) defined the older Pompeckium kuthani and the younger Bohemiella
romingeri biozones (Fig. 1.E ![]() ). Kordule
(2006) proposed a
problematic scheme combining three barren intervals with four assemblage zones
of rare ptychopariid and paradoxidid trilobites along with the earlier
established trilobite and brachiopod biozones.
). Kordule
(2006) proposed a
problematic scheme combining three barren intervals with four assemblage zones
of rare ptychopariid and paradoxidid trilobites along with the earlier
established trilobite and brachiopod biozones.
The discovery of the trilobite Mikaparia mutica (Hawle & Corda, 1847) (V. Vokáč, personal communication) suggests that the Biskoupky locality belongs to the middle to upper levels of the Buchava Formation (cf. Kordule, 2006; Vokáč & Micka, 2024). However, other faunal elements typically associated with younger strata of the Buchava Formation are absent. Therefore, the interpretation of the biostratigraphic position of the Biskoupky locality based on macrofossils within the sequence of the Buchava Formation is uncertain.
The first report on organic-walled microfossils from the Buchava Formation was published by Konzalová (1974, p. 47, 49). It was followed by a study of Vavrdová (1976, p. 61-62), who analysed samples from three outcrops: the Pod hruškou, Buchava, and Dlouhá hora localities; acritarchs from the Dlouhá hora locality were further discussed by Vavrdová (1982, p. 146-147). From stratigraphically high levels of the Skryje Shale at the Luh locality, acritarchs were reported by Chlupáč et al. (1998, p. 171, interval 6).
The reports generally describe moderately diversified assemblages consisting of Timofeevia sp., Solisphaeridium spp., small acanthomorphic acritarchs (Micrhystridium, Heliosphaeridium), and leiospheres. Furthermore, Chlupáč et al. (1998) demonstrated the presence of Eliasum llaniscum and Cristallinium cambriense - two taxa important in Miaolingian biostratigraphy.
A set of small outcrops is located approximately 850 m NW of the village of Biskoupky. These outcrops are dispersed across the middle portion of a slope above the Škaredá rivulet. The sequence represents an isolated relict of Miaolingian sediments, roughly 100 × 30 m in size, enclosed within grey-blue to purple andesites and rhyolites of the Křivoklát-Rokycany Volcanic Complex. At the Biskoupky locality, the sediments are tectonically bounded by faults, presumed to trend NW and NE (Kettner, 1923; Vorel et al., 2018). Cleavage overprints the original stratification substantially; however, measurements taken from the outcrops indicate a generally N-S, steeply inclined bedding orientation.
Massive greyish-green siltstones dominate the sequence, forming layers more than 30 cm thick. Within these siltstones, fine to medium-grained, sometimes laminated greywackes and easily disintegrating shales form subordinate layers. The sequence seems to exhibit a generally upward fining trend; however, minor layers of greywackes occasionally occur higher up in the section as well. In the central portion, carbonate-rich concretions are present.
The first fossils from the Biskoupky locality were reported by Knittl (1913). Subsequent discoveries of skeletal fauna and ichnofossils were reported by Smetana (1920), V. Kraft (1928), Šnajdr (1958), Havlíček (1977), J. Kraft (1982), Mergl and Šlehoferová (1990), Vokáč (1997), Mikuláš (2000), Kordule (2006), Mergl and Zamora (2013), and Nolčová and Mergl (2016). More recently, Valent et al. (2017, p. 229) briefly summarised all previously published data on skeletal fauna and listed the presence of trilobites (nine genera), agnostids (one species), lingulate brachiopods (two species), rhynchonelliformean brachiopods (one species), echinoderms (two species), helcionelloid molluscs (two species), hyoliths (seven species), indeterminable sponge spicules, and four genera of ichnofossils.
Six samples of fine-grained siltstone to shale (ca. 20-30 g each) were collected. All but one sample (BS-BI-0) were processed using a standard HCl-HF-HCl palynological procedure (e.g., Riding, 2021). Neither centrifugation nor oxidation were applied. Residues were passed through a 32µm mesh, and the smaller fraction was subsequently filtered through a 10µm mesh. BS-BI-0 was treated only with HF and sieved at 30µm. Part of the smaller fraction was subsequently treated with HCl and heated in order to remove secondary fluorides. The resulting residue was filtered through a 10µm mesh.
Microfossils in the > 32µm fraction were handpicked by pipette and mounted on slides, fixed by synthetic resin (Entellan), and covered with a cover slip. For the 10-32µm fraction, a drop of residue was mixed with glycerin gel, spread over a slide, and covered with a cover slip. Fossils were documented using an Olympus BX 51 optical microscope. Slides are deposited in the Czech Geological Survey under numbers VKO27 to VKO53.
The residues contain isolated acritarchs (see Figs. 2 ![]() - 3
- 3 ![]() ),
acritarch clusters (for comparison, see Kovář et al.,
2023) and a few cyanobacterial filaments. The preservation of individual
specimens varies considerably. While processes are generally well-preserved,
including multiple levels of branching, a considerable portion of the
processes is often broken off. This prevents a precise reconstruction of the
original number of processes for the individual specimens and taxa. Central
bodies of acritarchs are often plastically deformed due to early diagenetic
pyrite growth, making taxonomic determination of many specimens difficult (see
Fig.
4
),
acritarch clusters (for comparison, see Kovář et al.,
2023) and a few cyanobacterial filaments. The preservation of individual
specimens varies considerably. While processes are generally well-preserved,
including multiple levels of branching, a considerable portion of the
processes is often broken off. This prevents a precise reconstruction of the
original number of processes for the individual specimens and taxa. Central
bodies of acritarchs are often plastically deformed due to early diagenetic
pyrite growth, making taxonomic determination of many specimens difficult (see
Fig.
4 ![]() ). Curiously, specimens of Eliasum Fombella,
1977, emend. Palacios, 2015, appear
to be affected only rarely, which stands in sharp contrast with other taxa.
This suggests a biologically and/or morphologically selective pyrite formation
within different acritarchs.
). Curiously, specimens of Eliasum Fombella,
1977, emend. Palacios, 2015, appear
to be affected only rarely, which stands in sharp contrast with other taxa.
This suggests a biologically and/or morphologically selective pyrite formation
within different acritarchs.
In addition to the clearly Miaolingian-aged acritarchs, two
specimens of comparatively large, poorly preserved, degraded and highly
flattened acritarchs are preserved (see Fig. 4.J-K ![]() ). These specimens exhibit a clearly
distinct mode of preservation differing from the rest of the microfossil
assemblage. One of the specimens represents a leiosphere (Fig.
4.J
). These specimens exhibit a clearly
distinct mode of preservation differing from the rest of the microfossil
assemblage. One of the specimens represents a leiosphere (Fig.
4.J ![]() );
the other is comparable to the genus Germinosphaera Mikhailova,
1986, emend. Butterfield, 1994
(Fig. 4.K
);
the other is comparable to the genus Germinosphaera Mikhailova,
1986, emend. Butterfield, 1994
(Fig. 4.K ![]() ).
).
|
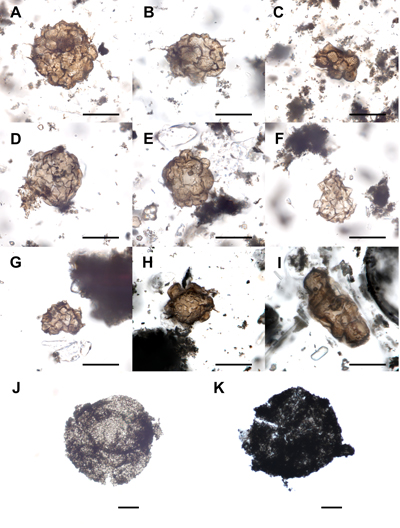
Figure 2: Specimens of acritarchs from the Biskoupky locality. A - Leiosphaeridia minutissima; B, D - Heliosphaeridium cf. notatum; C - Solisphaeridium sp.; E-F - Heliosphaeridium cf. exile; G - Heliosphaeridium sp.; H-I - Retisphaeridium dichamerum; K-L - Cristallinium cambriense; J, M - Eliasum llaniscum. The scale bar represents 20 µm. England finder (EF) coordinates for individual fossils on slides stored at the Czech Geological Survey (Prague): Specimen A: Slide BS-BI-1-Sc_10 EF:G53/4; |
|
Figure 3: Specimens of acritarchs from the Biskoupky locality. A - Adara? sp.; B - acritarch indet.; C - Timofeevia simplex; D-F - Timofeevia lancarae; G-H - Timofeevia heteromorpha; I - Timofeevia cf. raquelinae; J-K - clusters of Timofeevia. The scale bar represents 20 µm. England finder (EF) coordinates for individual fossils on slides stored at the Czech Geological Survey (Prague): Specimen A: Slide BS-BI-0-Sc_30 EF:H62/3; |
The sequence is primarily composed of siltstones typically lacking internal stratification, though poorly developed laminations may occasionally be observed. This locally develops into up to 1 cm-thick layers of shale. Fragments of irregularly dispersed remains of skeletal fossils occur in the silty sediment and generally are not concentrated in distinct fossiliferous horizons. The character of the sedimentation is thus consistent with distal turbidite deposition dominated by fine-grained material deposited in numerous separate pulses. A coarser fraction of the turbidites is represented by layers of mostly fine-grained greywackes up to several centimetres thick; these locally also preserve current-induced sedimentary structures typical for the upper portions of a Bouma sequence (Bouma, 1962). The greywackes are made up of quartz grains corresponding in size to the silty to fine sandy fraction, argillized rock clasts and comparatively rare heavy minerals such as zircons, apatite, baryte, rutile, and less frequently also leucoxene and tourmaline. The matrix is composed of clay minerals with secondary Fe minerals associated. The clays are represented mainly by highly crystalline illite, occasionally replaced by chlorite or chlorite aggregates. Despite the relict's position within the volcanic complex, the sediments are not substantially more thermally affected than is common elsewhere within the Skryje-Týřovice Basin.
Organic residues contain a substantial amount of amorphous organic
matter and, curiously, a considerable proportion of acritarchs are severely
deformed by pyrite ingrowths - both framboidal (e.g., Fig.
4.B-C ![]() )
and cubical (e.g., Fig. 4.F-G
)
and cubical (e.g., Fig. 4.F-G ![]() ) forms
- from inside. The
acritarch walls reacted with a high degree of plasticity, replicating both the
surface texture and the overall shape of the pyrite grains. This, alongside
the degree of crystallinity exhibited by the pyrite grains, suggests a
synsedimentary to early diagenetic origin for the pyrite at least partially
linked to the decay of organic matter within the acritarchs.
) forms
- from inside. The
acritarch walls reacted with a high degree of plasticity, replicating both the
surface texture and the overall shape of the pyrite grains. This, alongside
the degree of crystallinity exhibited by the pyrite grains, suggests a
synsedimentary to early diagenetic origin for the pyrite at least partially
linked to the decay of organic matter within the acritarchs.
|
Figure 4: Examples of reworked (J-K) and severely deformed acritarchs (A-I) from the Biskoupky locality. Specimens A, B, C, E are not assigned to any taxon; specimens D, F, G likely represent leiospheres. Specimen I likely belongs to the genus Eliasum, however, it could also be a closer undetermined fusiform acritarch. The scale bar represents 20 µm. England finder (EF) coordinates for individual fossils on slides stored at the Czech Geological Survey (Prague): Specimen A: Slide BS-BI-1-Sd_10 EF:N44/2; |
The synonymy lists are not exhaustive and generally only include a limited number of publications that are most important for understanding the taxonomy of the respective species or their occurrence within the Cambrian of the Skryje-Týřovice Basin.
Group Acritarchs Evitt, 1963 (incertae sedis)
Genus Adara Fombella, 1977,
emend. Martin in Martin & Dean,
1981
Type species. Adara matutina Fombella, 1977.
Adara? sp.
(Fig. 3.A ![]() )
)
Remarks: Only a single specimen tentatively assigned to the genus Adara
has been observed. The specimen measures 44.3 μm in diameter and bears
several tens of conical processes with wide bases and blunt tips, ranging from
wide cones to longer, distally more cylindrical processes. The size of the
vesicle and the morphology of processes generally resemble those of A.
longispinosa Fatka, 1989, which has been synonymized with A.
alea Martin in Martin & Dean,
1981, by Jachowicz-Zdanowska (2013);
however, there are substantially fewer processes in the specimen from
Biskoupky. The specimen further resembles an acritarch specimen figured by Palacios
et al. (2021, Fig. 25.B), identified there as Adara alea.
Numerous other specimens resemble Adara to a degree (e.g., Fig.
4.F-G ![]() ); however, these are in fact often specimens of different acritarchs,
likely mostly leiospheres, that have been severely deformed by pyrite.
); however, these are in fact often specimens of different acritarchs,
likely mostly leiospheres, that have been severely deformed by pyrite.
Genus Cristallinium Vanguestaine,
1978,
emend. Palacios in Palacios et al.,
2022
Type species. Cristallinium cambriense (Slavíková, 1968) Vanguestaine, 1978, emend. Palacios in Palacios et al., 2022 (= Dictyotidium cambriense Slavíková, 1968, p. 201, Pl. II, fig. 1).
Cristallinium cambriense (Slavíková,
1968) Vanguestaine, 1978,
emend. Palacios in Palacios
et al.,
2022
(Fig. 2.K-L ![]() )
)
1968 Dictyotidium cambriense n. sp. - Slavíková, p. 201, Pl. II, figs. 1, 3.
1978 Cristallinium cambriense (Slavíková, 1968) comb. nov. - Vanguestaine, p. 271, Pl. II, figs. 16-17; Pl. 3, figs. 16, 26.
1981 Cristallinium cambriense - Martin in Martin & Dean, p. 17, Pl. 3, figs. 4- 5, 9, 11; Pl. 5, figs. 3, 5, 8, 11.
1998 Cristallinium cambriense - Chlupáč et al., p. 171.
2022 Cristallinium cambriense emend. - Palacios et al., p. 32-33, Fig. 25.B.
Remarks: Only three observed specimens of polygonal acritarchs exhibit a well-developed minute ornamentation along their edges. Distinguishing whether this is the result of taphonomic loss of ornamentation or reflects a genuine rarity of Cristallinium is difficult to determine unequivocally. However, specimens of Cristallinium, while present, are generally not very abundant in the Cambrian deposits of both the Příbram-Jince and Skryje-Týřovice basins (Kovář, unpublished data). Specimens herein assigned to Cristallinium cambriense range between 30.6 and 43.7 µm (N=3).
Genus Eliasum Fombella,
1977,
emend. Palacios, 2015
Type species. Eliasum llaniscum Fombella, 1977.
Eliasum llaniscum Fombella, 1977
(Fig. 2.J, .M ![]() )
)
1977 Eliasum llaniscum n. sp. - Fombella, p. 118, Pl. I, fig. 6.
1998 Eliasum llaniscum - Chlupáč et al., p. 171.
1998 Eliasum llaniscum - Moczydłowska, p. 66-68, Fig. 28.A-D.
2015 Eliasum llaniscum - Palacios, Pl. VIII, fig. 1.
Remarks: Eliasum llaniscum is a common and easily
recognizable component of the microfossil assemblage at the Biskoupky
locality.
The specimens are mostly comparatively mildly deformed; severe pyrite
ingrowths occur only rarely. Complete specimens range from 33.1 to 75.8 µm
(N=41; μ=59.2 µm; σ=14.6) in length and lack surface ornamentation.
Apart from isolated specimens, three clusters consisting of two to three
specimens have been recovered. No other species of Eliasum has been
observed; however, a few poorly preserved specimens are assigned to the genus
only tentatively and could belong to an undetermined fusiform acritarch genus
instead, as these specimens lack discernible longitudinal ridges (e.g.,
Fig. 4.I ![]() ).
).
Genus Germinosphaera Mikhailova,
1986,
emend. Butterfield, 1994
Type species. Germinosphaera bispinosa Mikhailova, 1986.
Germinosphaera? sp.
(Fig. 4.K ![]() )
)
Remarks: A single, poorly preserved specimen measures approximately 98.5 µm in diameter. At least two processes are present; however, they are broken off near their bases. The width of the processes at their base corresponds to 11.8 and 12.8 µm, respectively. Its preservation is distinct from that of the Miaolingian acritarchs preserved in the sample, indicating it is possibly reworked material.
Genus Heliosphaeridium Moczydłowska, 1991
Type species. Heliosphaeridium dissimilare (Volkova, 1969) Moczydłowska, 1991.
Material: A total of 35 specimens were examined. However, the majority of specimens have all but a few processes broken off, and many specimens are damaged, severely limiting the possibility of species-level determination.
Heliosphaeridium cf. notatum (Volkova, 1969) Moczydłowska, 1991
(Fig. 2.B, .D ![]() )
)
cf. 1969 Micrhystridium notatum n.sp. - Volkova, p. 228, Pl. 51, figs. 16-19 (non vidi).
cf. 1991 Heliosphaeridium notatum (Volkova 1969) comb. nov. - Moczydłowska, p. 58.
cf. 1998 Heliosphaeridium notatum - Moczydłowska, p. 77-79, Figs. 31.L, 33.A-L.
cf. 2013 Ammonidium notatum comb. nov. - Jachowicz-Zdanowska, p. 47-48, Pl. XXII, figs. 13-15, 17-24.
Remarks: Subspherical to oval acritarchs with processes bearing funnel-shaped endings. In one specimen, short branching is observed in the distal parts of the processes. Central body size ranges from 12.8 to 16.8 µm (N=4). Considerable variability in process length is evident both among specimens and within a single specimen (cf. Moczydłowska, 1998); the processes range from 2 to 8.3 µm.
Heliosphaeridium cf. exile Moczydłowska, 1998
(Fig. 2.E-F ![]() )
)
cf. 1998 Heliosphaeridium exile sp. n. - Moczydłowska, p. 75, Pl. 32, fig. G.
Remarks: Subspherical to oval acritarchs with short processes that have acute tips and conical bases. The size of the central body ranges from 16.1 to 25.9 µm (N=5). Locally, the wall appears concave between processes, though this is difficult to ascertain due to deformations of the central body. Approximately 10-15 processes are visible on the observed surface; in contrast, H. lanceolatum (Vanguestaine, 1974) Moczydłowska, 1998, typically bears 20-40 processes.
Heliosphaeridium sp.
(Fig. 2.G ![]() )
)
Remarks: These are subspherical to oval acritarchs with slightly conical to hair-like processes ending in sharp tips. The central body size ranges from 14.8 to 23.4 µm (N=5), and process lengths are variable, ranging from 2.4 to 8.1 µm.
Genus Leiosphaeridia Eisenack, 1958
Type species. Leiosphaeridia baltica Eisenack, 1958.
Leiosphaeridia minutissima Naumova,
1949,
emend. Jankauskas in Jankauskas et al.,
1989
(Fig. 2.A ![]() )
)
1949 Leiotriletes minutissimus sp. n. - Naumova, p. 52-53, Tab. 1, figs. 1-2, Tab. 2, figs. 1-2.
1989 Leiosphaeridia minutissima (Naumova, 1949) emend., comb. nov. - Jankauskas et al., p. 79-80, Pl. IX, figs. 1-4, 11.
Remarks: Numerous specimens can be assigned to Leiosphaeridia
with various certainty levels. The diameter ranges from 12.5 to 65.7 µm
(N=64; μ=29.8 µm; σ=10.6). These specimens are often deformed by
pyrite ingrowths, which, in some cases, causes them to resemble other genera,
especially Adara (see, e.g., Fig. 4.G ![]() ). A similar
deformation is also common in specimens of other genera (e.g., Solisphaeridium
or Timofeevia) in our material. In some specimens, the severe
deformation prevents an unequivocal differentiation between Leiosphaeridia
and Retisphaeridium.
). A similar
deformation is also common in specimens of other genera (e.g., Solisphaeridium
or Timofeevia) in our material. In some specimens, the severe
deformation prevents an unequivocal differentiation between Leiosphaeridia
and Retisphaeridium.
Leiosphaeridia sp.
(Fig. 4.J ![]() )
)
Remarks: A single, poorly preserved specimen of Leiosphaeridia. The specimen is oval in outline (likely deformed), measuring approximately 108.6 µm in maximal length. The preservation suggests that the specimen is reworked, as is the case with the specimen of Germinosphaera? sp.
Genus Retisphaeridium Staplin et al.,
1965,
emend. Palacios
in Palacios et al.,
2022
Type species. Retisphaeridium dichamerum Staplin et al., 1965, emend. Palacios in Palacios et al., 2022.
Retisphaeridium dichamerum Staplin et al.,
1965,
emend. Palacios
in Palacios et al.,
2022
(Fig. 2.H-I ![]() )
)
1965 Retisphaeridium dichamerum sp. n. - Staplin et al., p. 187-188, Pl. 19, figs. 1-7.
1982 Retisphaeridium dichamerum - Downie, p. 279, Figs. 11 o-p.
2022 Retisphaeridium dichamerum Staplin et al., 1965, emend. - Palacios et al., p. 36, Figs. 13.A-F.
Remarks: A range of taphomorphs ranging from specimens clearly exhibiting their original subspherical form to those resembling Cymatiosphaera (see Downie, 1982; Palacios et al., 2022). Complete specimens range from 12.7 to 50.7 µm (N=31; μ=33.6; σ=7.99) in diameter and have approximately 13-18 fields on the observed side. However, the exact number of fields is difficult to ascertain in many specimens due to their deformation and/or degradation. Further spherical to subspherical acritarch specimens, severely deformed by pyrite, could also belong to the genus; however, these specimens could also be placed into Leiosphaeridia or other sphaeromorph taxa.
Genus Solisphaeridium Staplin et al.,
1965,
emend. Moczydłowska, 1998
Type species Solisphaeridium stimuliferum (Deflandre, 1938) Staplin et al., 1965.
Solisphaeridium spp.
(Fig. 2.C ![]() )
)
Remarks: Originally spherical acritarchs with processes. The size of the central body ranges from 24.7 to 52.7 µm (N=36). The processes are primarily slender, cylindrical to hairlike, though more rarely conical, reaching up to 18.9 µm. The majority of processes are broken off, which hampers more precise determination. However, most of our specimens are generally comparable to Solisphaeridium flexipilosum Slavíková, 1968.
Genus Synsphaeridium Eisenack, 1965
Type species. Synsphaeridium gotlandicum Eisenack, 1965.
Synsphaeridium? sp.
Remarks: Two clusters of closely packed leiosphere-like objects consisting of three and six spheres, respectively. The individual spheres are approximately 10 µm in diameter.
Genus Timofeevia Vanguestaine, 1978
Type species. Multiplicisphaeridium lancarae Cramer & Diéz, 1972.
Remarks: As with other acritarchs in the studied material, a large portion of the specimens is substantially degraded and/or affected by pyrite ingrowths, obscuring the original outline of the central body. Curiously, processes tend to be comparatively well-preserved, exhibiting up to several levels of branching. Timofeevia occurs both individually and in clusters. We follow the species classification developed by Palacios (2015) and Palacios et al. (2022), though a revision of the species co-occurring in the T. lancarae Zone is warranted.
Timofeevia heteromorpha Palacios, 2015
(Fig. 3.G-H ![]() )
)
2015 Timofeevia heteromorpha sp. nov. - Palacios, p. 94-96, Pl. XI, figs. 1-6.
2022 Timofeevia heteromorpha - Palacios et al., Fig. 26.F.
Remarks: Polygonal to subspherical acritarchs, unless secondarily deformed by pyrite. The size of the central body ranges from 18.6 to 46 µm (N=20). Processes conical to subcylindrical, ranging from simple to branched in up to three orders. Process length varies from 3.1 to 13.5 µm.
Timofeevia lancarae (Cramer & Diéz, 1972) Vanguestaine, 1978
(Fig. 3.D-F ![]() )
)
1972 Multiplicisphaeridium lancarae sp. n. - Cramer & Diéz, p. 42, Pl. 1, figs 1-4, 6, 8.
1978 Timofeevia lancarae (Cramer & Diéz) nov. comb. - Vanguestaine, p. 272.
2015 Timofeevia lancarae - Palacios, p. 96-98, Pl. XII, figs. 1-6.
Remarks: Polygonal to subspherical acritarchs, unless secondarily deformed by pyrite. The size of the central body ranges from 17.8 to 47.8 µm (N=42; μ=30.6 µm; σ=6.5). Processes conical, often with broad bases. Processes usually branch several times in their distal portions, forming pinae: process length is variable, ranging from 3.6 to 13.5 µm.
Timofeevia cf. raquelinae (Cramer & Díez, 1972) Cramer & Diéz, 1979
(Fig. 3.I ![]() )
)
cf. 1972 Multiplicisphaeridium raquelinae (New Species). - Cramer & Díez, p. 43-44, Pl. 1, fig. 7.
cf. 1979 Timofeevia raquelinae - Cramer & Díez, p. 58.
Remarks: Polygonal to subspherical acritarchs, unless secondarily deformed by pyrite. The size of the central body ranges from 26.9 to 39.8 µm (N=4). Processes conical to subcylindrical, with pointed or manate endings, rarely extending into short branches. Process length varies from 3.6 to 9.8 µm. The specimens are similar to those figured by Palacios (2015). The presence of T. raquelinae has also been reported by Vavrdová (1982); however, no specimens were figured.
Timofeevia simplex Palacios, 2015
(Fig. 3.C ![]() )
)
2015 Timofeevia simplex sp. n. - Palacios, p. 98, Pl. XIII, figs. 1-4.
Remarks: Polygonal to subspherical acritarchs, unless secondarily deformed by pyrite. The size of the central body ranges from 24 to 41.5 µm (N=14). Processes conical or hair-like with conical bases and pointed tips, and only rarely do the tips branch dichotomously. The process length varies considerably from 3.7 to 10.8 µm.
Apart from Miaolingian acritarchs, two reworked acritarchs were documented. These
can be distinguished by their very poor, highly flattened preservation and the
comparatively darker colouration of the wall. One of the reworked acritarchs
likely represents a large leiosphere (Fig. 4.J ![]() ), while the second
specimen is tentatively assigned to the genus Germinosphaera (Fig.
4.K
), while the second
specimen is tentatively assigned to the genus Germinosphaera (Fig.
4.K ![]() ). Comparable fossils have been reported from Proterozoic sequences of
the Teplá-Barrandian Unit (e.g., Fatka & Gabriel,
1991).
). Comparable fossils have been reported from Proterozoic sequences of
the Teplá-Barrandian Unit (e.g., Fatka & Gabriel,
1991).
The reworked acritarchs and analyses of heavy minerals indicate that the source material is derived from weakly metamorphosed sedimentary and magmatic rocks of the Cadomian basement. This is in accordance with the data of Drost et al. (2004). The sedimentary rocks incorporated into the Cadomian basement span a prolonged timespan, demonstrated to include fossiliferous sequences of both Proterozoic and Cambrian age (see Konzalová, 1974; Hajná et al., 2017). Both Proterozoic and "early" Cambrian Cadomian rocks lie relatively close to the Skryje-Týřovice Basin, making them likely sources for the clastic material within the basin. Unfortunately, the current stratigraphic utility of the reworked microfossils is quite limited. Nevertheless, their presence suggests that at least part of the clastic material at the Biskoupky locality more likely originates from the Neoproterozoic rather than the Cambrian sequences. Additional material could prove useful for clarifying the provenance of sediments within the Příbram-Jince and Skryje-Týřovice basins.
The acritarch record from the Biskoupky locality consists of a distinct assemblage including abundant specimens of Eliasum llaniscum, Retisphaeridium dichamerum, multiple species of Timofeevia and Heliosphaeridium, as well as Solisphaeridium sp. and rare specimens of Cristallinium cambriense. Our findings generally agree with records from other localities within the Skryje-Týřovice Basin (Konzalová, 1974; Vavrdová, 1976, 1982; Chlupáč et al., 1998; Kovář & Fatka, 2024). Noteworthy is the presence of Adara?; Adara is commonly found across European peri-Gondwana and Baltica (see summary in Palacios et al., 2022) and is notably present in the Příbram-Jince Basin of the TBU (Fatka, 1989; Kovář & Fatka, 2025). However, Adara has not previously been observed in the Skryje-Týřovice Basin (see Section 2.2 and Kovář & Fatka, 2024). Whether the conspicuous lack of Adara in other successions is due to the age of the successions, unknown ecological or paleobiogeographic constraints, or insufficient sampling cannot be determined at this time. Unfortunately the single specimen is insufficient for any further interpretations.
In the Příbram-Jince Basin, Timofeevia first appears within the Onymagnostus hybridus Biozone (see Fatka, 1989; Kovář et al., 2023; Kovář & Fatka, 2025). However, based on the macrofossil content, the rocks of the Skryje-Týřovice Basin have been compared to the P. (Eccaparadoxides) pussilus Biozone, which partly underlies the O. hybridus Biozone in the Jince Formation with an only relatively short overlap. The acritarch record, therefore, suggests that the P. (Eccaparadoxides) pussilus biozones of the Příbram-Jince and Skryje-Týřovice basins might not be directly correlatable and that the studied sections within the Skryje-Týřovice Basin are likely younger than previously expected (cf. e.g., Šnajdr, 1958; Havlíček, 1971; Kordule, 2006). More generally, the acritarch assemblage at the Biskoupky locality closely resembles those of the T. lancarae Zone described by Palacios et al. (2022), which has been suggested to span the late Drumian to early Guzhangian. If accepted, this age range would contradict the age indicated by the trilobites due to the occurrence of "Paradoxides" hicksii in the Skryje-Týřovice Basin (see Vaněk et al., 1999), a trilobite species characteristic of the lower half of the Drumian (compare Fletcher, 2007; Geyer, 2019). Further analyses of both macrofossil and microfossil records of the Skryje-Týřovice Basin is necessary in order to explain these discrepancies.
(1) Organic-walled microfossils are reported from the southernmost occurrence of the Buchava Formation in the Skryje-Týřovice Basin for the first time. The acritarchs include, most notably, Eliasum llaniscum, Retisphaeridium dichamerum, Solisphaeridium sp., multiple species of Timofeevia and Heliosphaeridium, and rare specimens of Cristallinium cambriense. Notably, Adara? sp. is described for the first time from the basin.
(2) The assemblage closely matches with assemblages of the Timofeevia lancarae Zone described by Palacios et al. (2022) and its analogues in peri-Gondwana and Baltica.
(3) The stratigraphic interpretation based on the microfossil record differs slightly from the biostratigraphy based on macrofossils (especially trilobites).
We thank Marika Polechová for translating the abstract into French. We are very grateful to the editor Bruno Granier and both reviewers, Marco Vecoli and an anonymous reviewer, for their commentary, constructive criticism, and suggestions, which substantially improved the manuscript. We thank Zev Brook for assistance with language editing. This research was supported by the Grant Agency of Charles University, project no. 264523 (V. Kovář), and by Cooperatio - Geol of the Ministry of Education, Youth and Sports of the Czech Republic (O. Fatka). This manuscript contributes to the Strategic Research Plan of the Czech Geological Survey (DKRVO/ČGS 2023-2027). This is a contribution to the IGCP project 735 'Rocks and the Rise of Ordovician Life' (2021-2025).
Barrande J. (1846).- Notice préliminaire sur le Systčme Silurien et les Trilobites de Bohęme.- Hirschfeld, Leipsic, 97 p.
Barrande J. (1852).- Systčme Silurien du Centre de la Bohęme, Ičre Partie, Crustacés, Trilobites.- 2 volumes, chez l'auteur et éditeur, Prague, Paris, 935 p. URL: https://archive.org/details/systemesilurien1barr
Bouma A.H. (1962).- Sedimentology of some flysch deposits: A graphic approach to facies interpretation.- Elsevier, Amsterdam, 168 p.
Bruthansová J., Fatka O., Budil P. & Král J. (2007).- 200 years of trilobite research in the Czech Republic. In: Mikulic M.G., Landing E. & Kluessendorf J. (eds.), Fabulous fossils - 300 years of worldwide research on trilobites.- New York State Museum Bulletins, New York - NY, vol. 507, p. 51-80.
Butterfield N.J., Knoll A.H. & Swett K. (1994).- Paleobiology of the Neoproterozoic Svanbergfjellet Formation, Spitsbergen.- Fossils and Strata, Oslo, No. 34, p. 1-84. DOI: 10.1111/j.1502-3931.1994.tb01558.x
Chlupáč I. (1999).- Barrande's stratigraphic concepts, palaeontological localities and tradition - comparison with the present state.- Journal of the Czech Geological Society, Prague, vol. 44, nos. 1-2, p. 3-30.
Chlupáč I., Fatka O., Prokop R.J. & Turek V. (1998).- Výzkum klasické paleontologické lokality "Luh" ve skryjském kambriu (Research of the classical paleontological locality "Luh" in the Cambrian of Skryje).- Journal of the Czech Geological Society, Prague, vol. 43, no. 3, p. 169-173.
Colthurst J.R.J. & Smith D.G. (1977).- Palaontological evidence for the age of the Lower Palaeozoic rocks of the Slievenamon Inlier, County Tipperary.- Proceedings of the Royal Irish Academy (Section B: Biological, Geological, and Chemical Science), Dublin, vol. 77, p. 143-158.
Cramer F.H. & Diéz M.B.C.R. (1972).- Acritarchs from the Upper Middle Cambrian Oville Formation of Leon, Northwestern Spain.- Revista Espańola de Micropaleontologia, Madrid, Numero extraordinario, p. 39-50.
Cramer F.H. & Diéz M.B.C.R. (1979).- Lower Paleozoic acritarchs [Acritarcos del Paleozoico Inferior].- Instituto de Investigaciones Palinológicas, Léon (Spain), p. 17-160.
Deflandre G. (1938).- Microplancton des mers jurassiques conservé dans les marnes de Villers-sur-Mer (Calvados). Étude liminaire et considérations générales.- Travaux de la Station Zoologique de Wimereux, Paris, vol. 13, p. 147-200.
Downie C. (1982).- Lower Cambrian acritarchs from Scotland, Norway, Greenland and Canada.- Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 72, no. 4, p. 257-285.
Drost K., Linnemann U., Mcnaughton N., Fatka O., Kraft P., Gemlich M., Tonk C. & Marek J. (2004).- New data on the Neoproterozoic-Cambrian geotectonic setting of the Teplá-Barrandian volcano-sedimentary successions: Geochemistry, U-Pb zircon ages, and provenance (Bohemian Massif, Czech Republic).- International Journal of Earth Sciences, vol. 93, no. 5, p. 742-757.
Eisenack A. (1958).- Tasmanites Newton 1875 und Leiosphaeridia n.g. als Gattungen der Hystrichosphaeridea.- Palaeontographica Abteilung A, Stuttgart, vol. 110, nos. 1-3, p. 1-19.
Eisenack A. (1965).- Mikrofossilien aus dem Silur Gotlands Hystichosphären, Problematika.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 122, no. 3, p. 257-274.
Evitt W.R. (1963).- A discussion and proposals concerning fossil dinoflagellates, hystrichospheres, and acritarchs, II.- Proceedings of the National Academy of Sciences of the United States of America, Washington - DC, vol. 49, no. 3, p. 298-302.
Fatka O. (1989).- Acritarch assemblage in the Onymagnostus hybridus Zone (Jince Formation, middle Cambrian, Czechoslovakia).- Věstník Ústředního ústavu geologického, Prague, vol. 64, no. 6, p. 363-367.
Fatka O. (1990).- Das Kambrium von Skryje und Týřovice. In: Weidert K.H. (ed.), Klassische Fundstellen der Paläontologie, Band 2.- Goldschneck-Verlage, Stuttgart, p. 12-17.
Fatka O. (2004).- Association of fossils and history of research at the Týřovice - Pod hruškou locality (Middle Cambrian, Skryje-Týřovice Basin, Barrandian area).- Journal of the Czech Geological Society, Prague, vol. 49, no. 3, p. 107-117.
Fatka O. & Gabriel Z. (1991).- Microbiota from siliceous stromatolitic rocks of the Barrandian Proterozoic (Bohemian Massif).- Časopis pro mineralogii a geologii, Prague, vol. 36, nos. 2-3, p. 143-148.
Fatka O. & Herynk J. (2016).- The first occurrence of bivalved arthropod Tuzoia from the Skryje-Týřovice Basin (Barrandian area, Czech Republic).- Annales de Paléontologie, Paris, vol. 102, no. 4, p. 219-224.
Fatka O., Kraft P. & Szabad M. (2011a).- Shallow-Water Occurrence of Wiwaxia in the Middle Cambrian of the Barrandian Area, Czech Republic.- Acta Palaeontologica Polonica, Warsaw, vol. 56, no. 4, p. 871-875.
Fatka O., Micka V., Szabad M., Vokáč V. & Vorel T. (2011b).- Nomenclature of Cambrian lithostratigraphy of the Skryje-Týřovice Basin.- Bulletin of Geosciences, Prague, vol. 85, no. 4, p. 841-858. URL: http://www.geology.cz/bulletin/contents/art1284
Fletcher T.P. (2007).- Correlating the zones of 'Paradoxides hicksii' and 'Paradoxides davidis' in Cambrian Series 3.- Memoirs of the Association of Australasian Palaeontologists, Canberra, vol. 33, p. 35-56.
Fombella M.A. (1977).- Acritarcos de edad Cámbrico Medio-Inferior de la provincia de León, Espańa.- Revista Espańola de Micropaleontología, Madrid, vol. 9, no. 1, p. 115-124.
Geyer G. (2019).- A comprehensive Cambrian correlation chart.- Episodes, Seoul, vol. 42, no. 4, p. 321-332. DOI: 10.18814/epiiugs/2019/019026
Geyer G., Elicki O., Fatka O. & Żylińska A. (2008).- Cambrian. In: McCann T. (ed.), Geology of Central Europe.- Geological Society of London, London, p. 155-202.
Hajná J., Žák J. & Dörr W. (2017).- Time scales and mechanisms of growth of active margins of Gondwana: A model based on detrital zircon ages from the Neoproterozoic to Cambrian Blovice accretionary complex, Bohemian Massif.- Gondwana Research, Beijing, vol. 42, p. 63-83.
Havlíček V. (1971).- Stratigraphy of the Cambrian of Central Bohemia.- Sborník geologických Věd, Geologie, Prague, vol. 20, p. 7-52.
Havlíček V. (1977).- Brachiopods of the order Orthida in Czechoslovakia.- Rozpravy Ústředního Ústavu geologického, Prague, vol. 44, 327 p.
Hawle I. & Corda A.J.C. (1847).- Prodrom einer Monographie der böhmischen Trilobiten.- Calve, Prague, 176 p.
Jachowicz-Zdanowska M. (2013).- Cambrian phytoplankton of the Brunovistulicum - taxonomy and biostratigraphy.- Polish Geological Institute Special Papers, Warsaw, vol. 28, p. 1-150.
Jahn J.J. (1896).- Ueber die geologischen Verhältnisse des Kambrium von Tejřovic und Skrej in Böhmen.- Jahrbuch der Kaiserlichen-königlichen Geologischen Reichsanstalt, Wien, vol. 45, p. 641-790.
Jankauskas T.V., Mikhailova N.S. & Hermann T.N. (eds., 1989).- Mikrofossilii Dokembriya SSSR.- Nauka, Leningrad, 190 p.
Kettner R. (1923).- Kambrium skrejsko-tejřovické a jeho okolí [Cambrian of the Skryje-Týřovice area and its surroundings].- Sborník Státního geologického ústavu Československé republiky, Prague, vol. 3, p. 5-63 [in Czech].
Knittl Č. (1913).- O útvaru kamenouhelném u Vejvanova [On the Carboniferous at Vejvanov].- Brdský kraj, Rokycany, vol. 5, p. 150-153 [in Czech].
Konzalová M. (1974).- Acritarchs from the Bohemian Precambrian (Upper Proterozoic) and Lower-Middle Cambrian.- Review of Palaeobotany and Palynology, vol. 18, nos. 1-2, p. 41-56.
Kordule V. (2006).- Ptychopariid trilobites in the Middle Cambrian of Central Bohemia (taxonomy, biostratigraphy, synecology).- Bulletin of Geosciences, Prague, vol. 81, no. 4, p. 277-304. URL: http://www.geology.cz/bulletin/contents/art2006.04.277
Kovář V. & Fatka O. (2024).- Acritarch assemblages of the Skryje-Týřovice Basin and their biostratigraphic implications. In: Soták J., Kyška Pipík R. & Tomašových A. (eds.), 23rd Czech-Slovak-Polish Paleontological Conference: Abstract book.- Earth Science Institute, Slovak Academy of Science, Bratislava, p. 48.
Kovář V. & Fatka O. (2025).- The first record of Hallucigenia-like lobopodians from the lower Jince Formation (Cambrian, Miaolingian) of the Příbram-Jince Basin.- Swiss Journal of Paleontology, London, vol. 144, no. 38, p. 1-13.
Kovář V., Fatka O. & Vodička J. (2023).- Acritarch clusters from the Cambrian (Miaolingian) of the Příbram-Jince Basin, Czech Republic.- Palynology, Abingdon, vol. 47, no. 1, p. 1-18.
Kraft J. (1982).- Chráněná a k ochraně navržená paleontologická naleziště na Rokycansku Rokycany.- Sborník Západočeského muzea v Plzni, Příroda, Pilsen, vol. 45, p. 1-51 [in Czech].
Kraft V. (1928).- Geologické poměry Rokycanska [Geology of the Rokycany area].- Rokycany, 121 p. [in Czech].
Lipold M.V. & Krejčí J. (1860).- Verhandlungen der Kaiserlichen-königlichen geologischen Reichsanstalt, Wien, vol. 11, p. 88-91.
Maletz J., Steiner M. & Fatka O. (2005).- Middle Cambrian pterobranchs and the Question: What is a graptolite?- Lethaia, Oslo, vol. 38, no. 1, p. 73-85.
Martin F. & Dean W.T. (1981).- Middle and Upper Cambrian and Lower Ordovician acritarchs from Random Island, Eastern Newfoundland.- Geological Survey of Canada Bulletin, Ottawa, vol. 343, p. 1-43.
Mergl M. & Šlehoferová P. (1990).- Middle Cambrian inarticulate brachiopods from Central Bohemia.- Sborník geologických Věd, Paleontologie, Prague, vol. 31, p. 67-104.
Mergl M. & Zamora S. (2013).- New and revised occurrences of rhynchonelliformean brachiopods from the middle Cambrian of the Iberian Chains, NE Spain.- Bulletin of Geosciences, Prague, vol. 87, no. 3, p. 571-586. URL: http://www.geology.cz/bulletin/contents/art1327
Mikhailova N.S. (1986).- Novye nakhodki mikrofitofossilij iz otlozhenijverkhnego rifeya Krasnoyarskogo kraya [New finds of microphytofossils from upper Riphean deposits of the Krasnoyar region]. In: Sokolov B.S. (ed.), Aktual'nye Voprosy Sovremennoj Paleoalgologii.- Naukova Dumka, Kyiv, p. 31-37.
Mikuláš R. (2000).- Trace fossils from the Middle Cambrian of the Barrandian area (Central Bohemia, Czech Republic).- Czech Geological Survey Special Papers, Prague, vol. 12, p. 1-29.
Mikuláš R. & Fatka O. (2017).- Ichnogenus Astropolichnus in Cambrian of the Barrandian area, Czech Republic.- Ichnos, Abingdon, vol. 24, no. 4, p. 283-290. DOI: 10.18261/8200374742-1991-01
Moczydłowska M. (1991).- Acritarch biostratigraphy of the Lower Cambrian and the Precambrian-Cambrian boundary in southeastern Poland.- Fossils and Strata, Oslo, vol. 29, 127 p. DOI: 10.18261/8200374742-1991-01
Moczydłowska M. (1998).- Cambrian acritarchs from Upper Silesia, Poland - biochronology and tectonic implications.- Fossils and Strata, Oslo, vol. 46, 121 p. DOI: 10.18261/8200376923-1998-01
Naumova S.N. (1949).- Spory nizhnego kembriya [Spores from the Lower Cambrian].- Izvestiya Akademii Nauk SSSR, Seriya Geologicheskaya, Moscow, vol. 1949, no. 4, p. 49-56.
Nolčová L. & Mergl M. (2016).- Výskyt epibiontních ostnokožců v buchavském souvrství (kambrium, drum) u Biskoupek na Radnicku (Barrandien) [Occurrence of epibiont echinoderms in the Buchava Formation (Cambrian, Drumian) at Biskoupky near Radnice (Barrandian area, Czech Republic)].- Zprávy o geologických výzkumech, Prague, vol. 49, p. 43-46 [in Czech with English summary].
Palacios T. (2015).- Acritarch assemblages from the Oville and Barrios Formations, northern Spain: A pilot proposal of a middle Cambrian (Series 3) acritarch biozonation in northwestern Gondwana.- Review of Palaeobotany and Palynology, vol. 219, p. 71-105.
Palacios T., Jensen S., Álvaro J.J., Santos Zaldeugui J.F., Eguiluz L, Corfu F. & Gil Ibarguchi J.I. (2021).- Acritarch-based chronostratigraphic and radiometric calibration of the Cambrian volcanosedimentary Vallehondo and Playón formations in the Cambrian Ossa-Morena Rift, Spain.- Palæogeography, Palæoclimatology, Palæoecology, vol. 565, article 110216, 32 p.
Palacios T., Högström A.E.S., Jensen S., Ebbestad J.O.R., Agić H., Hřyberget M., Meinhold G. & Taylor W. (2022).- Organic-walled microfossils from the Kistedalen Formation, Norway: Acritarch chronostratigraphy of the Baltic Miaolingian and evolutionary trends of placoid acritarchs.- Papers in Palaeontology, Durham, vol. 8, no. 4, article e1457, 41 p. DOI: 10.1002/spp2.1457
Pompeckj J.F. (1896).- Die Fauna des Kambrium von Tejřovic und Skrej in Böhmen.- Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, Wien, vol. 45, p. 495-614.
Riding J.B. (2021).- A guide to preparation protocols in palynology.- Palynology, Abingdon, vol. 45, no. S1, p. 1−110.
Slavíková K. (1968).- New finds of acritarchs in the Middle Cambrian of the Barrandian (Czechoslovakia).- Věstník Ústředního ústavu geologického, Prague, vol. 43, no. 3, p. 199-205.
Smetana V. (1920).- Kambrium u Biskoupek na Zbirožsku [Cambrian in the Zbiroh area].- Rozpravy II. třídy České akademie, Prague, vol. 28, no. 23, p. 1-4 [in Czech].
Staplin F.L., Jansonius J. & Pocock S.A.J. (1965).- Evaluation of some acritarchous hystrichosphere genera.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 123, no. 2, p. 167-201.
Šnajdr M. (1958).- Trilobiti českého středního kambria (Trilobites of the Bohemian Middle Cambrian).- Rozpravy Ústředního ústavu geologického, Prague, vol. 26, 280 p. [in Czech with English summary].
Vaněk J., Valíček J. & Vokáč V. (1999).- Plutonides hicksi (Salter) from the Middle Cambrian of Skryje-Týřovice area (Czech Republic).- Palaeontologia Bohemiae, Prague, vol. 5, no. 6, p. 36-38.
Valent M., Fatka O. & Marek L. (2017).- Biskolites iactans gen. et sp. nov. from the Cambrian of the Czech Republic (Hyolitha, Skryje-Týřovice Basin).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 285, no. 2, p. 227-233.
Vanguestaine M. (1974).- Espčces zonales d'Acritarches du Cambro-Trémadocien de Belgique et de l'Ardenne française.- Review of Palaeobotany and Palynology, vol. 18, p. 63-82.
Vanguestaine M. (1978).- Critčres palynostratigraphiques conduisant ŕ la reconnaissance d'un pli couché Revinien dans le sondage de Grand-Halleux.- Annales de la Société géologique de Belgique, Ličge, vol. 100, p. 249-276. URL: https://popups.uliege.be/0037-9395/index.php?id=4694
Vavrdová M. (1976).- Excystment mechanism of Early Paleozoic acritarchs.- Časopis pro mineralogii a geologii, Prague vol. 21, no. 1, p. 55-64.
Vavrdová M. (1982).- Phytoplankton communities of Cambrian and Ordovician age of Central Bohemia.- Věstník Ústředního ústavu geologického, Prague, vol. 57, no. 3, p. 145-155.
Vokáč V. (1997).- Middle Cambrian benthic assemblage in the Skryje Shale (Jince Formation) at Terešovská Huť (district of Rokycany, Czech Republic).- Palaeontologia Bohemiae, Prague, vol. 3, p. 15-19 [in Czech with English summary].
Vokáč V. (2002).- Poznámky k biostratigrafii jineckého souvrství (střední kambrium) v skryjsko-týřovickém regionu (Barrandien, Čechy). [Remarks on the biostratigraphy of the Jince Formation (Middle Cambrian) from Skryje-Týřovice region (Barrandian, Bohemia)].- Palaeontologia Bohemiae, Prague, vol. 8, no. 6, p. 117-118 [in Czech with English summary].
Vokáč V. & Micka V. (2024).- Diverzita trilobitových asociací několika lokalit buchavského souvrství (kambrium, drum) skryjsko-týřovické pánve, Barrandien, Česká republika [Diversity of the trilobite assemblages of several localites in the Buchava Formation, (Cambrian, Drumian) in the Skryje-Týřovice Basin, Barrandian, Czech Republic].- Sborník Západočeského muzea v Plzni - Příroda, Pilsen, vol. 130, p. 1−89 [in Czech with English summary].
Volkova N.A. (1969).- Akritarkhi severo-zapada Russkoj platformy [Acritarchs of the north-western Russian platform]. In: Rozanov A.Yu., Missarzhevsky V.V., Volkova N.A., Voronova L.G., Krylov I.N., Keller B.M., Korolyuk I.K., Lendzion K., Michniak R., Pykhova N.G. & Sidorov A.D. (eds.), Tommotskij yarus i problema nizhnej granitsy kembriya [Tommotian Stage and the Cambrian lower boundary problem].- Nauka, Moscow, p. 224−236.
Vorel T., Dušek K., Havlín Nováková D., Holásek O., Hradecký P., Hrazdíra P., Janderková J., Kotková J., Krupička J., Mašek D., Sidorinová T., Skácelová D., Skácelová Z., Soejono I., Stárková M. & Zelenková Trubačová A. (2018).- Základní geologická mapa České republiky 1:25 000, list 12-323 Podmokly [Basic geological map of the Czech Republic 1:25 000, sheet 12-323 Podmokly].- Czech Geological Survey Prague [in Czech].
Table of reports on acritarchs from the Skryje-Týřovice Basin with acritarch taxa reported.
| Konzalová (1974, p. 47, 49) |
|
| Vavrdová (1976, p. 61-62) |
|
| Vavrdová (1982, p. 146-147) |
|
| Chlupáč et al. (1998, p. 171) |