◄ Carnets Geol. 26 (7) ►
![]()
Outline:
[1. Introduction]
[2. The restricted geometries of abiotic processes]
[3. The expanded geometries of biological systems]
[4. The narrow overlap ...]
[5. Empirical support for a morphogeometric theory ...]
[6. Implications for astrobiology ...]
[7. Toward a general morphogeometric theory of life]
[8. Conclusions]
and ... [Bibliographic references]
Institute
of Ecology and Earth Sciences, University of Tartu, Ravila 14A, 50411, Tartu (Estonia)
Published online in final form (pdf) on May 21, 2026
DOI
10.2110/carnets.2026.2607
![]()
[Editor:
Robert W. Scott; technical editor: Bruno Granier]
![]()
Current evolutionary theory provides detailed accounts of biological diversity, but it still lacks a framework capable of describing the geometric-topological structures in a unified way that living systems produce. Physical and chemical processes can generate only a limited variety of geometric forms, whereas biological morphogenesis explores a much larger and more complex region of morphospace. This distinction has important implications for paleobiology, the biology, and astrobiology: if life produces geometries unavailable to physical-chemical non-living processes, then geometry itself may serve as a universal biosignature. I argue that developing a general morphogeometric theory of life is both conceptually necessary and practically valuable for identifying life in the geological record and beyond Earth.
morphogeometry;
biosignatures;
morphospace;
biogenicity assessment;
abiotic-biotic discrimination;
fossil identification algorithms
Vinn O. (2026).- Toward a universal geometry of life: Implications for biological theory and astrobiology.- Carnets Geol., Madrid, vol. 26, no. 7, p. 151-161. DOI: 10.2110/carnets.2026.2607
Vers une géométrie universelle du vivant : Implications pour la théorie biologique et l'astrobiologie.- La théorie actuelle de l'évolution fournit des descriptions détaillées de la diversité biologique, mais elle ne dispose pas encore d'un cadre capable de décrire, de manière unifiée, les structures géométrico-topologiques produites par les systèmes vivants. Les processus physiques et chimiques ne peuvent générer qu'une variété limitée de formes géométriques, tandis que la morphogenèse biologique explore une région du morphoespace beaucoup plus vaste et complexe. Cette distinction a des implications importantes pour la paléobiologie, la biologie et l'astrobiologie : si la vie produit des géométries inaccessibles aux processus physico-chimiques non vivants, alors la géométrie elle-même pourrait servir de biosignature universelle. Je soutiens que le développement d'une théorie morphogéométrique générale du vivant est à la fois conceptuellement nécessaire et pratiquement précieuse pour identifier la vie dans les archives géologiques et au-delà de la Terre dans l'espace.
morphogéométrie ;
biosignatures ;
morphoespace ;
évaluation de la
biogénicité ;
discrimination abiotique/biotique ;
algorithmes
d'identification des fossiles
Since the publication of D'Arcy
Thompson's "On Growth and Form", researchers have recognized
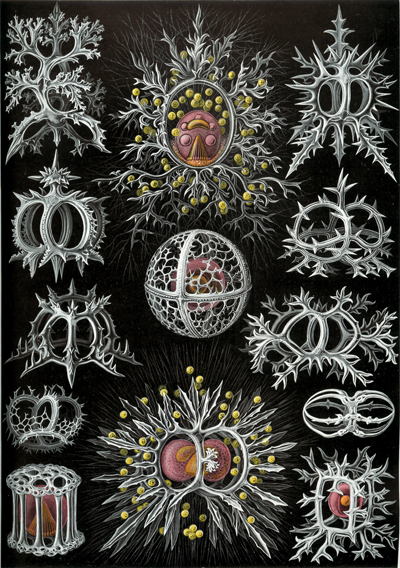
that biological shapes (Fig. 1 ![]() )
reflect mathematical and physical principles,
yet these insights remain fragmented (Thompson, 1917/1942). Modern
biology describes development, evolution, and biomechanics in increasingly
sophisticated terms, but it still lacks an overarching theoretical account of
how living systems generate form as such, despite important contributions from
theoretical morphology, constructional morphology, and morphometrics (Seilacher,
1970, 1973, 1979,
1989, 1991; Vogel,
1991; McGhee, 1999; Mitteroecker
& Huttegger, 2009; Budd,
2021). What is missing is a general
morphogeometric theory that describes the geometric domains accessible to life,
the mechanisms that produce biological form, and the distinctions between
biogenic and abiogenic structures, building on classic notions of morphospace
and their philosophical interpretation (Raup, 1966; Kaandorp,
2008; Polly, 2023). Such a theory would illuminate the geometry of life
across different substrates and environments, complementing existing
evolutionary and developmental frameworks (Gould, 2002; Müller
& Newman, 2003).
)
reflect mathematical and physical principles,
yet these insights remain fragmented (Thompson, 1917/1942). Modern
biology describes development, evolution, and biomechanics in increasingly
sophisticated terms, but it still lacks an overarching theoretical account of
how living systems generate form as such, despite important contributions from
theoretical morphology, constructional morphology, and morphometrics (Seilacher,
1970, 1973, 1979,
1989, 1991; Vogel,
1991; McGhee, 1999; Mitteroecker
& Huttegger, 2009; Budd,
2021). What is missing is a general
morphogeometric theory that describes the geometric domains accessible to life,
the mechanisms that produce biological form, and the distinctions between
biogenic and abiogenic structures, building on classic notions of morphospace
and their philosophical interpretation (Raup, 1966; Kaandorp,
2008; Polly, 2023). Such a theory would illuminate the geometry of life
across different substrates and environments, complementing existing
evolutionary and developmental frameworks (Gould, 2002; Müller
& Newman, 2003).
An additional perspective can be gained by considering the broader intellectual landscape represented in the work of Douglas H. Erwin (2007, 2008, 2015) and David Jablonski (2007, 2008, 2017). Their research situates biological form within deep evolutionary time, emphasizing that morphology is not only shaped by immediate functional demands but also by historical contingency, developmental constraints, and large-scale evolutionary dynamics. This body of work draws on paleobiology, evolutionary developmental biology (evo-devo), and macroevolutionary theory to show how new forms arise, persist, and diversify. In particular, it highlights that the emergence of novel geometries - such as new body plans or structural architectures - cannot be understood solely in terms of local physical processes but must be seen as the outcome of evolving developmental programs and ecological interactions.
A major problem that Precambrian paleontologists often encounter, and astrobiologists always face, is pseudofossils (McMahon, 2020). These are naturally occurring, non-biological structures that can be confused with genuine fossils because they resemble biological shapes. Certain mineral growth patterns are especially deceptive, since inorganic processes can produce forms that look unexpectedly intricate or organized. A well-known case is the dendritic pattern formed by crystallizing manganese oxides, which spread through rock fractures in branching, tree-like arrangements that superficially imitate fossilized plants. To definitively refute the abiogenic origin of structures, we need, in addition to chemical-isotopic composition, also working geometric-morphological criteria.
Here, I propose a hypothesis that structures produced by the abiotic and biotic domains mainly have distinct geometries with only minor overlap among the simplest geometric forms, and that a general mathematical geometrical theory can, in most cases, tell us the origin of structures without any doubt. The aim of this paper is to discuss the feasibility and necessity of a morphogeometric theory, and its potential astrobiological and paleobiological implications. The morphogeometric theory is defined here as a theory capable of distinguishing biological structures from abiotic structures solely on the basis of their geometric properties.
|
|
Figure 1:
Radiolarians by Ernst Haeckel - Kunstformen der Natur (1904), Plate 71:
Stephoidea. |
Abiotic processes can produce visually striking structures such as crystals, fractal dendrites, layered chemical deposits, and reaction-diffusion patterns; nevertheless, these forms belong to a relatively narrow and well-understood range of mathematical possibilities (Turing, 1952; Kauffman, 1993). Their shapes typically arise from symmetry constraints, energy minimization, and simple reaction-diffusion dynamics, which restrict their geometric diversity. Detailed studies of mineral self-organization and silica-carbonate biomorphs have shown that nonliving systems can mimic certain simple biological forms, but only within a constrained set of morphologies and growth regimes (García-Ruiz et al., 2003, 2020; Rouillard et al., 2018; Cuéllar-Cruz, 2021). Although such structures may superficially resemble biological ones, they generally lack internal differentiation, hierarchical organization, and the capacity for adaptive or directed growth; their morphologies are fixed by environmental conditions, not by historically evolved programs or functional demands (García-Ruiz et al., 2017; Board on Life Sciences, 2018). Internal differentiation refers here to the presence of distinct parts within a structure that perform different roles. In living systems, form is not uniform: different regions are specialized, whether at the level of tissues in an organism or functional zones in a microbial colony. For example, a plant stem contains outer protective layers, internal transport tissues, and supportive structures, each with a specific function. By contrast, a diffusion-limited aggregate or a mineral dendrite is compositionally and functionally uniform throughout; it may be complex in shape, but it does not divide into specialized regions. Hierarchical organization means that a structure is arranged across multiple, clearly distinguishable levels, where smaller components are nested within larger ones in a systematic way. In biology, this often takes the form of branching systems organized into orders: large trunks divide into smaller branches, which divide further into fine branches, each level serving a related but distinct role. The human lung or a tree's root system are typical examples. In contrast, although abiotic structures like diffusion-limited aggregation clusters can be fractal and branching, they lack clear functional levels; their complexity is scale repeating rather than organized into discrete tiers with differentiated roles. Diffusion-Limited Aggregation (DLA) is a canonical stochastic growth model introduced by Thomas A. Witten Jr and Leonard M. Sander (1981) to describe structures formed when particles undergoing diffusion irreversibly stick upon contact. Adaptive or directed growth refers to the ability of a system to modify its form in response to environmental conditions in a way that improves its performance or persistence. Living systems do not grow blindly: roots extend toward water, shoots toward light, and vascular networks reorganize to maintain efficient transport when conditions change. This involves feedback between the organism and its environment. By comparison, structures formed by purely physical processes such as diffusion-limited aggregation grow according to fixed rules without feedback or correction; they cannot redirect growth in response to changing conditions or functional demands.
In conclusion, limited morphogeometric repertoire sets a boundary on what nonliving matter can accomplish through physical-chemical processes alone.
Biological processes are far more complex than any known natural abiotic processes, allowing living structures to create geometries that can never be matched by simpler abiotic processes. Some architectural examples of biological systems uniquely exhibit: 1. Curvature fields with correlation and regulation, 2. Branching networks obeying optimization-scaling laws, 3. Nontrivial cycle structure (anastomosis) with functional redundancy, and 4. Tight coupling between geometry and flow.
Living structures are not just curved - they are curved in a coordinated and controlled way. The bends, folds, and shapes you see in an organism are not random; nearby parts tend to curve in related ways because underlying biological processes regulates growth. A leaf does not wrinkle arbitrarily. Its surface curvature is shaped by controlled growth differences between tissues, producing smooth, continuous forms like gentle bending or systematic rippling. Similarly, blood vessels curve gradually and coherently rather than forming sharp, irregular kinks.
When biological systems branch, they do so in a way that balances efficiency - typically minimizing energy use while maximizing transport (of fluids, nutrients, or signals). The sizes of branches are not random; they follow consistent proportional relationships. In the circulatory system, a large blood vessel divides into smaller ones in such a way that blood flow remains efficient. The diameters of the daughter vessels are related to those of the parent vessel to minimize resistance. Similar patterns appear in tree branches and root systems, where the structure supports both transport and mechanical stability.
Nontrivial cycle structure (i.e., anastomosis) with functional redundancy means that, when biological networks contain loops, multiple pathways that connect the same points. This creates redundancy: if one path is blocked or damaged, flow can be rerouted through another. In many animals, blood vessels form interconnected networks rather than simple trees. If one vessel becomes blocked, blood can still reach tissues through alternate routes. Fungal networks and leaf vein systems also show this kind of looping structure, which improves resilience.
Tight coupling between geometry and flow means that in living systems, shape and function are closely linked. The geometry of a structure (i.e., its size, curvature, and arrangement) is directly related to how something moves through it - such as fluids, nutrients, or signals - and the system can adjust its form accordingly. In the lungs, the branching pattern and size of airways are matched to airflow, ensuring that oxygen is delivered efficiently to all regions. If conditions change (e.g., sustained stress or damage), biological systems can remodel - altering vessel thickness or connectivity - to maintain effective flow.
The only abiotic structures that can match the complexity of living structures are those produced by artificial technological processes. Biological structures combine both complex biochemical processes and biological evolution, allowing them to explore geometric regions that are effectively inaccessible to natural abiotic processes. Living forms commonly exhibit hierarchical layering, functional optimization, and modular organization, such that shapes at one scale depend on and integrate with shapes at another, as seen in vascular networks, skeletal architectures, and shell morphologies (Raup, 1966; McGhee, 1999; Jablonski, 2008). Such characteristics would allow coding for distinguishing biotic and abiotic structures. Hierarchical layering means that a biological structure is built in nested levels, where larger structures are composed of smaller substructures, which are themselves composed of even smaller elements. Each level has its own role but also contributes to the whole. Functional optimization means here that biological shapes are not arbitrary - they are tuned to perform specific tasks efficiently, often balancing competing demands such as strength, transport, and energy use. Modular organization refers here to the construction of a system from repeating or semi-independent units (modules) that can function on their own but also work together as part of a larger system. This allows flexibility, repair, and adaptability. Developmental processes impose constraints that generate families of related but variably expressed forms, producing distinctive clusters within morphospace rather than isolated shapes (Mitteroecker & Huttegger, 2009; Gerber, 2017). Because these structures arise from sustained thermodynamic disequilibrium, inheritance, and adaptive feedback, they display geometric complexity that cannot be generated by chemistry or physics alone, as emphasized by work on evolutionary novelties and developmental programs (Müller & Newman, 2003; Wagner, 2014; Erwin, 2015). Biological geometry, therefore, reflects not only physical constraints but also the action of selection on variational properties over deep time (Gould, 1989; Kauffman, 1993). Living systems exist in a constant state of being out of equilibrium. They continuously take in energy (e.g., sunlight, chemical energy) and use it to build and maintain structure. In nonliving systems, structures typically form as energy is minimized; crystals grow into regular, repetitive shapes and then stop. In contrast, living systems keep growing, reorganizing, and maintaining complex forms because energy is constantly flowing through them.
The reason most geometries of living organisms differ from non-biological ones is that non-biological geometries are generally functionally inadequate for biological needs. While such geometries may occasionally suffice for the simplest forms of life, they do not meet the requirements of more complex biological systems.
Some simple biological forms - such as filaments, spheres, or laminated domes - can be mimicked by chemical and geological processes, and this overlap gives rise to long-standing controversies over certain microfossils and stromatolitic structures (Cady et al., 2003; Westall et al., 2015). Silica-carbonate biomorphs, framboidal pyrite, and dendritic mineral growths are now well documented as abiotic analogues that can complicate biogenicity assessments, yet they occupy only a small region of potential morphogeometric space and lack multiscale hierarchy or functional organization (García-Ruiz et al., 2003; Rouillard et al., 2018; Tenelanda-Osorio et al., 2025). The vast majority of biological geometry involves features like branching across multiple scales, orderly segmentation, directional curvature adapted to mechanical or physiological tasks, and modular repetition with variation, none of which arise naturally in nonliving systems as far as current empirical studies show (Callefo et al., 2019; Shkolyar et al., 2025). These characteristics would be amenable to coding that would distinguish biotic and abiotic structures. This explains why a small subset of possible fossils continues to be contentious, while most fossils, including trace fossils, are unmistakably biological: the morphological distance between complex biogenic structures and their closest abiotic analogues is substantial and systematically structured rather than accidental.
Biological structures often branch repeatedly, and this branching happens at several levels - from large divisions down to very fine structures. The same general pattern, a main structure divided into smaller parts, is repeated, but each level has its own size and role. A tree branches from the trunk to large branches, then to small branches, and at last to twigs. Similarly, in the human body, arteries branch into smaller vessels, which eventually lead to tiny capillaries. This multi-scale branching allows efficient resource distribution throughout the entire system. Orderly segmentation means that a structure is divided into repeating units arranged in a regular, organized sequence. These segments are not random - they follow a consistent pattern and often have similar shapes or functions. The vertebrate spine is made of a series of vertebrae arranged in order. Annelids and many arthropods have bodies divided into repeating segments. In plants, stems are organized into repeating units (nodes and internodes) where leaves and branches emerge. This segmentation provides both flexibility and structural organization. Directional curvature refers here to the fact that biological structures tend to bend or grow in specific, non-random directions, often guided by internal programs or external cues. Plant shoots bend toward light (i.e., phototropism), and roots curve downward or toward water sources. Animal structures also show directional shaping - for instance, the curvature of bones or shells is not arbitrary but follows growth patterns that support their function.
The main empirical support for
the morphogeometric theory comes from paleontology, where biological origin is
frequently inferred from form rather than material composition. Fossil remains
are often mineralogically indistinguishable from the surrounding rock matrix,
having undergone complete chemical replacement during diagenesis (Anderson
et al., 2023). This is particularly
true for many Palaeozoic fossils, and to a lesser extent, for Mesozoic and
Cenozoic fossils, in which original organic material has been entirely replaced
by silica, calcite, or other minerals (Anderson et al.,
2023). Despite this compositional
equivalence, such
structures are confidently identified as fossils on the basis of their geometry
(Figs. 2 ![]() - 3
- 3 ![]() ), including
symmetry, repetition, hierarchical organization, and
functional morphology. Paleontologists routinely rely only on geometric
reasoning when determining whether a mineral structure is biogenic, even if this
reliance is rarely formalized explicitly. Professional paleontologists usually
find it easy to distinguish pseudofossils from actual fossils, though the
corresponding geometrical analysis is carried out without seeing its
mathematical background. The organization of chambers in foraminifera, spiral
shells of gastropods and nautiloids, or the patterned repetition found in
segmented and modular organisms are all diagnostic evidence of underlying
developmental programs and functional constraints, not as spontaneous mineral
growth (Jablonski, 2008; Erwin,
2008; Kaandorp, 2008).
Moreover, larger-scale geometries containing smaller-scale geometries are
characteristic of well-preserved fossils. The well-preserved brachiopod and
mollusk shells exhibit, in addition to external morphology, also internal finer
scale structures, growth lamellae, which themselves are composed of biologically
controlled crystals forming the finest structural level. Usually, the geometry
of fossilized organisms differs from that of geological structures at different
scales, from macroscopic to microscopic, and even at the ultrastructural level.
The hierarchical geometry of a mineral structure is not, in itself, a sign of a
biogenic origin, but the specific architecture of each structural level
unambiguously testifies to a biological origin. Such geometric signatures are
treated as evidence for life because they reveal generative processes like
growth, behavior, and adaptation that cannot be accounted for by geochemical
self-organization alone, a point that underlies current methodological
frameworks for assessing biogenicity in deep time (Callefo et
al., 2019; Board on Life Sciences, 2018). Geometric reasoning is the common
way to determine that fossils of extinct groups with problematic systematic
positions are the remains of organisms rather than aberrant geological
sedimentary structures. Similarly, geometry is the basis for distinguishing
trace fossils (Fig. 4
), including
symmetry, repetition, hierarchical organization, and
functional morphology. Paleontologists routinely rely only on geometric
reasoning when determining whether a mineral structure is biogenic, even if this
reliance is rarely formalized explicitly. Professional paleontologists usually
find it easy to distinguish pseudofossils from actual fossils, though the
corresponding geometrical analysis is carried out without seeing its
mathematical background. The organization of chambers in foraminifera, spiral
shells of gastropods and nautiloids, or the patterned repetition found in
segmented and modular organisms are all diagnostic evidence of underlying
developmental programs and functional constraints, not as spontaneous mineral
growth (Jablonski, 2008; Erwin,
2008; Kaandorp, 2008).
Moreover, larger-scale geometries containing smaller-scale geometries are
characteristic of well-preserved fossils. The well-preserved brachiopod and
mollusk shells exhibit, in addition to external morphology, also internal finer
scale structures, growth lamellae, which themselves are composed of biologically
controlled crystals forming the finest structural level. Usually, the geometry
of fossilized organisms differs from that of geological structures at different
scales, from macroscopic to microscopic, and even at the ultrastructural level.
The hierarchical geometry of a mineral structure is not, in itself, a sign of a
biogenic origin, but the specific architecture of each structural level
unambiguously testifies to a biological origin. Such geometric signatures are
treated as evidence for life because they reveal generative processes like
growth, behavior, and adaptation that cannot be accounted for by geochemical
self-organization alone, a point that underlies current methodological
frameworks for assessing biogenicity in deep time (Callefo et
al., 2019; Board on Life Sciences, 2018). Geometric reasoning is the common
way to determine that fossils of extinct groups with problematic systematic
positions are the remains of organisms rather than aberrant geological
sedimentary structures. Similarly, geometry is the basis for distinguishing
trace fossils (Fig. 4 ![]() )
from abiogenic sedimentological structures (Crimes
& Droser, 1992). There is only a tiny overlap between the morphology
of sedimentary structures and trace fossils. Thus, empirically, not only the
geometry of organisms but also the geometry of traces left by living organisms
are usually distinct from geological structures.
)
from abiogenic sedimentological structures (Crimes
& Droser, 1992). There is only a tiny overlap between the morphology
of sedimentary structures and trace fossils. Thus, empirically, not only the
geometry of organisms but also the geometry of traces left by living organisms
are usually distinct from geological structures.
A formal morphogeometric theory would make these ideas more explicit by identifying which kinds of shapes can only be produced by living systems, and by clarifying the basis on which scientists decide whether a structure, such as a fossil, a pseudo-fossil, or a microbially induced sedimentary structure was formed by biological activity (Gerber, 2017; Polly, 2023). In doing so, it would help resolve borderline cases where morphology is suggestive but not conclusive, and it would integrate geometric evidence with chemical and contextual data into a more coherent evidential framework.
|
|
Figure 2:
Trilobite Asaphus (GIT 438-404) from
the Ordovician of Russia exhibiting fossilized exoskeleton and undoubtedly
biotic geometry. Collection of Tallinn University of Technology, Department of
Geology. Photo by Gennadi Baranov. |
|
|
Figure 3:
Fish Leptolepis (TUG 400-12) from Upper Jurassic of Solnhofen, Germany.
Fossilized skeleton and undoubtedly biotic geometry. Collection of Natural
History Museum, University of Tartu. Scale bar 1 cm. Photo by Mare Isakar. |
|
|
Figure 4:
Palaeozoic Zoophycos.
Trace fossils exhibiting internal structure (modified after Seilacher,
2007, Pl. 37). |
Contemporary astrobiological approaches often prioritize chemical biosignatures and atmospheric disequilibria, yet chemistry-based reasoning becomes fragile when considering life with unknown or non-Earth-like biochemistries (Des Marais et al., 2003; National Academies of Sciences, Engineering, and Medicine, 2019). Geometry, by contrast, depends on generative processes such as differentiation, growth, and adaptive feedback that may be universal to evolving systems regardless whether it is supported by biochemistry similar to that of life on earth or an alien alternative, a point increasingly emphasized in recent discussions of structural or morphological biosignatures (Cady et al., 2003; Westall et al., 2015, 2021; Shkolyar et al., 2025). If evolving systems naturally explore complex regions of morphospace, then alien life forms should exhibit geometric structures that differ systematically from abiotic formations even when their chemistry is unfamiliar, and this expectation is already implicit in recommendations for future Mars and icy-moon missions (Des Marais et al., 2008; Westall et al., 2015; Tenelanda-Osorio et al., 2025). Geometric analysis could therefore allow the recognition of extraterrestrial fossils or biological traces even when their molecular composition has degraded or cannot be readily interpreted, provided that we have an explicit theory specifying which morphogeometric features are strongly diagnostic of life, such as branching across multiple scales, orderly segmentation, and directional curvature. Such a theory would complement, rather than replace, chemical and isotopic approaches by offering a chemistry-independent criterion grounded in the organization and complexity of form.
Morphogeometric theory would enable the design of algorithms capable of recognizing the distinctive structural features produced by living systems. Such a theory would specify the geometric signatures that arise from biological growth, differentiation, and functional adaptation, allowing these patterns to be distinguished from the more limited forms generated by abiotic processes. With these principles formalized, algorithmic tools could be trained not merely on examples from Earth but on abstract geometric rules that any life-like system is expected to follow. This would greatly reduce the reliance on Earth-specific biological analogues and expand our ability to identify more unfamiliar forms. Onboard robotic missions could then use these algorithms to scan high-resolution images for multiscale hierarchical organization, modularity, and non-random curvature, all of which are characteristic of living structures. Instead of blindly searching for familiar fossils, the software could evaluate whether a structure lies within the region of morphospace accessible to biological processes. The algorithms could also assess how well an observed pattern fits known physical or chemical self-organization models, thereby ruling out many pseudofossils. By integrating both positive biological criteria and negative abiotic constraints, such systems could make real-time decisions about which samples or locations warrant closer examination. This would dramatically increase the efficiency of life-detection missions on Mars, Europa, Enceladus, or exoplanetary surfaces. The approach would also help avoid misinterpretations by providing a rigorous geometric basis for rejecting ambiguous but ultimately inorganic structures. Because geometric principles of life may be universal, this method would not depend on detecting specific Earth-based chemistry. Instead, it would rely on the more general physical consequences of life's organizational complexity. By doing so, it would approach the problem of life detection from a broader theoretical perspective. Such tools could also play a role in selecting specimens for sample-return missions, ensuring that only the most promising materials are brought back to Earth. The algorithms might even help guide autonomous drilling or excavation systems toward sites with high biological potential. The underlying morphogeometric theory would also enable cross-comparison between different planetary environments, revealing whether similar life-like forms emerge under different conditions. In effect, these algorithms would serve as an intelligent filter between raw imagery and scientific interpretation. Ultimately, a morphogeometric theory of life would transform robotic photographic exploration from a passive data-collection exercise into an active, theory-guided search for signs of life.
In 2019, entomologist William S. Romoser (Ohio University) presented a conference poster titled "Does Insect/Arthropod Biodiversity Extend Beyond Earth?" at the Entomological Society of America meeting. His central claim was that publicly available NASA Mars rover images (especially those from Curiosity) contain visual evidence of 1) Insect-like organisms (with wings, segmented bodies, legs), 2) Reptile-/snake-like forms, 3) Both fossilized and living organisms, and 4) Apparent ecological interactions (e.g., predator-prey relationships). Romoser argued that recognizable anatomical traits (e.g., "exoskeleton," "jointed appendages," "wings") are sufficient to classify these as arthropod-like life forms beyond Earth (Romoser, 2019; Johnson, 2019). The Mars "insect" claims fail primarily because the data modality (rover imagery) is fundamentally inadequate for the level of biological inference being made. These data do not have sufficient spatial resolution and scale calibration. Interpretations unsupported by quantitative morphometric analysis are best explained by pareidolia (imposition of imagined meaningful interpretations on chance patterns) rather than by evidence of extraterrestrial arthropods. Future image interpretation algorithms using morphogeometric theory will be capable of refuting of confirming such claims.
A mature morphogeometric theory would map the geometric domains accessible to living versus nonliving systems and formulate the geometric principles underlying biological shape, drawing from the long tradition of theoretical morphology and recent advances in morphospace analysis (Raup, 1966; Seilacher 1970, 1979, 1991; McGhee, 1999; Mitteroecker & Huttegger, 2009; Gerber, 2017; Budd, 2021). It would describe how developmental constraints guide organismal form, how evolutionary processes populate morphospace over time, and how non-equilibrium dynamics enable geometries unavailable to passive physical systems, integrating insights from pattern-formation theory and complex systems science (Turing, 1952; Kauffman, 1993; Müller & Newman, 2003). The theory would also need to identify topological invariants of life - such as hierarchy, modularity, and functional integration - and explain how these features arise from universal organizational principles rather than from specific biochemistries, aligning with recent work on evolutionary innovation and constraint (Wagner, 2014; Erwin, 2015). The concept of theoretical morphospace already points in this direction by distinguishing the merely possible from the historically realized, but it has not yet been extended to systematically include abiotic morphogenesis and astrobiological contexts (McGhee, 1999; Kaandorp, 2008; Polly, 2023). Such a framework would unify insights from paleobiology, evolutionary theory, morphogenesis, and astrobiology into a single conceptual system that treats geometry as a central feature of life rather than as a decorative outcome.
There is massive data that needs to be collected before one can produce the morphogeometric theory of life. Data collection should begin with eigenshape data from a large number of organisms, both living and fossil, across various branches of the evolutionary tree. In addition, eigenshape data of geological structures should be collected to enable comparison. Creation of 3D models of organisms using CT scanning should be carried out on a large scale. Then AI can be trained to mathematically analyze massive eigenshape data to map the geometries of life and, if possible, to suggest mathematical generalizations and rules for geometric discrimination of biotic and abiotic structures.
Biology contains extensive empirical knowledge of form, yet it lacks a unified philosophical and theoretical account of how living systems generate geometric complexity and how this complexity differs, in principle, from nonliving structures. A general morphogeometric theory would clarify how biological and abiotic forms diverge, grounding paleobiological and astrobiological inferences on a more secure conceptual foundation and making explicit the assumptions already guiding the interpretation of fossils and biosignatures (Board on Life Sciences, 2018; Westall et al., 2021). Such a theory would also shift the search for life away from chemistry alone and toward the deeper principles governing organization, growth, and structural possibility, thereby opening new avenues for recognizing life that does not share Earth's biochemical details. As exploration extends to ancient terrestrial rocks and extraterrestrial environments, and as studies of mineral self-organization refine our understanding of abiotic complexity, the ability to identify life through its geometry becomes an increasingly pressing scientific and philosophical challenge (García-Ruiz et al., 2020; Shkolyar et al., 2025; Tenelanda-Osorio et al., 2025).
This research was supported by the Estonian Research Council grant PRG2591. I am grateful to Carlton E. Brett and two anonymous reviewers for the constructive comments on the manuscript.
Anderson R.P., Woltz C.R., Tosca N.J., Porter S.M. & Briggs D.E.G. (2023).- Fossilisation processes and our reading of animal antiquity.- Trends in Ecology and Evolution, vol. 38, no. 11, p. 1060-1071. DOI: 10.1016/j.tree.2023.05.014
Board on Life Sciences, Division on Earth and Life Studies, National Academies of Sciences, Engineering, and Medicine (2018).- An astrobiology strategy for the search for life in the universe.- The National Academies Press, Washington - DC, 188 p.
Briggs D.E.G. (2005).- Seilacher on the science of form and function. In: Evolving form and function: Fossils and development. Proceedings of a symposium honoring Adolf Seilacher for his contributions to paleontology, in celebration of his 80th birthday.- Peabody Museum of Natural History Yale University, New Haven - CT, 288 p.
Budd G.E. (2021).- Morphospace.- Current Biology, vol. 31, no. 19, p. R1181-R1185. DOI: 10.1016/j.cub.2021.08.040
Cady S.L., Farmer J.D., Grotzinger J.P., Schopf J.W. & Steele A. (2003).- Morphological biosignatures and the search for life on Mars.- Astrobiology, vol. 3, no. 2, p. 351-368.
Callefo F., Maldanis L., Teixeira V.C., Oliveira Abans R.A. de, Monfredini T., Rodrigues F. & Galante D. (2019).- Evaluating biogenicity on the geological record with synchrotron-based techniques.- Frontiers in Microbiology, vol. 10, 12 p. DOI: 10.3389/fmicb.2019.02358
Crimes T.P. & Droser M.L. (1992).- Trace fossils and bioturbation: The other fossil record.- Annual Review of Ecology and Systematics, vol. 23, p. 339-360.
Cuéllar-Cruz M. (2021).- Influence of abiotic factors in the chemical origin of life: Biomorphs as a study model.- ACS Omega, Washington - DC, vol. 6, no. 13, p. 8754-8763. DOI: 10.1021/acsomega.1c00497
Des Marais D.J., Allamandola L.J., Benner S.A., Boss A.P., Deamer D., Falkowski P.G., Farmer J.D., Hedges S.B., Jakosky B.M., Knoll A.H., Liskowsky D.R., Meadows V.S., Meyer M.A., Pilcher C.B., Nealson K.H., Spormann A.M., Trent J.D., Turner W.W., Woolf N.J. & Yorke H.W. (2003).- The NASA Astrobiology Roadmap.- Astrobiology, vol. 3, no. 2, p. 219-235.
Des Marais D.J., Nuth J.A. 3rd, Allamandola L.J., Boss A.P., Farmer J.D., Hoehler T.M., Jakosky B.M., Meadows V.S., Pohorille A., Runnegar B. & Spormann A.M. (2008).- The NASA Astrobiology Roadmap.- Astrobiology, vol. 8, no. 4, p. 715-730.
Erwin D.H. (2007).- Disparity: Morphological pattern and developmental context.- Paleontology, vol. 50, no. 1, p. 57-73. DOI: 10.1111/j.1475-4983.2006.00614.x
Erwin D.H. (2008).- Extinction as the loss of evolutionary history.- Proceedings of the National Academy of Sciences, vol. 105, Suppl. 1, p. 11520-11527. DOI: 10.1073/pnas.0801913105
Erwin D.H. (2015).- A public goods approach to major evolutionary innovations.- Geobiology, vol. 13, no. 4, p. 308-315.
García-Ruiz J.M., Hyde S.T., Carnerup A.M., Christy A.G.., Van Kranendonk M.J. & Welham N.J. (2003).- Self-assembled silica-carbonate structures and detection of ancient microfossils.- Science, vol. 302, no. 5648, p. 1194-1197.
García-Ruiz J.M., Nakouzi E., Kotopoulou E., Tamborrino L. & Steinbock O. (2017).- Biomimetic mineral self-organization from silica-rich spring waters.- Science Advances, vol. 3, no. 3, 6 p. DOI: 10.1126/sciadv.1602285
García-Ruiz J.M., Zuilen M.A. van & Bach W. (2020).- Mineral self-organization on a lifeless planet.- Physical of Life Reviews, vol. 34-35, p. 62-82.
Gerber S. (2017).- The geometry of morphospaces: Lessons from the classic Raup shell coiling model.- Biological Reviews, vol. 92, no. 2, p. 1142-1155.
Gould S.J. (1989).- Wonderful life: The Burgess Shale and the nature of history.- Norton, New York - NY, 352 p.
Gould S.J. (2002).- The structure of evolutionary theory.- Harvard University Press, Cambridge - MA, 1433 p. URL: https://archive.org/details/jaygouldthestructureofevolutionarytheory
Haeckel E. (1904).- Kunstformen der Natur.- Bibliographisches Institut, Leipzig, Wien, 73 p. (100 Pls.). URL: https://archive.org/details/KunstformenderN00Haec/page/n229/mode/2up
Jablonski D. (2007).- Scale and hierarchy in macroevolution.- Palaeontology, vol. 50, no. 1, p. 87-109. DOI: 10.1111/j.1475-4983.2006.00615.x
Jablonski D. (2008).- Biotic interactions and macroevolution: Extensions and mismatches across scales and levels.- Evolution, St. Louis - MO, vol. 62, no. 4, p. 715-739. DOI: 10.1111/j.1558-5646.2008.00317.x
Jablonski D. (2017).- Approaches to macroevolution: 1. General concepts and origin of variation.- Evolutionary Biology, vol. 44, no. 4, p. 427-450. DOI: 10.1007/s11692-017-9420-0
Johnson S. (2019).- Insects on Mars? Ohio scientist claims NASA images show life on red planet.- Big Think. URL: https://bigthink.com/hard-science/mars-aliens/
Kaandorp J.A. (2008).- Exploring theoretical morphospaces.- BioScience, Oxford, vol. 58, no. 3, p. 271-272.
Kauffman S.A. (1993).- The origins of order: Self-organization and selection in evolution.- Oxford University Press, Oxford, 729 p.
McGhee G.R. (1999).- Theoretical morphology: The concept and its applications.- Columbia University Press, New York - NY, 316 p.
McMahon S. (2020).- Earth's earliest and deepest purported fossils may be iron-mineralized chemical gardens.- Proceedings of the Royal Society B, vol. 286, article 20192410, 9 p. DOI: 10.1098/rspb.2019.2410
Mitteroecker P. & Huttegger S.M. (2009).- The concept of morphospaces in evolutionary and developmental biology: Mathematics and metaphors.- Biological Theory, vol. 4, no. 1, p. 54-67.
Müller G.B. & Newman S.A. (eds., 2003).- Origination of organismal form: Beyond the gene in developmental and evolutionary biology.- Vienna Series in Theoretical Biology, MIT Press, Cambridge - MA, 342 p.
National Academies of Sciences, Engineering, and Medicine (2019).- An astrobiology strategy for the search for Life in the universe.- The National Academies Press, Washington - DC, 184 p.
Polly P.D. (2023).- Extinction and morphospace occupation: A critical review.- Cambridge Prisms: Extinction, vol. 1, article e17, 9 p. DOI: 10.1017/ext.2023.16
Raup D.M. (1966).- Geometric analysis of shell coiling: General problems.- Journal of Paleontology, Tulsa - OK, vol. 40, no. 5, p. 1178-1190.
Romoser W.S. (2019).- Does insect/arthropod biodiversity extend beyond Earth? Poster presented at the Entomological Society of America meeting. DOI: 10.13140/rg.2.2.12363.95520 fatcat:deicfu4wifhktivjes2n7dqebi
Rouillard J., García-Ruiz J.M., Gong J. & Zuilen M.A. van (2018).- A morphogram for silica-witherite biomorphs and its application to microfossil identification in the early earth rock record.- Geobiology, vol. 16, no. 3, p. 279-296. DOI: 10.1111/gbi.12278
Seilacher A. (1970).- Arbeitskonzept zur Konstruktions-Morphologie.- Lethaia, Oslo, vol. 3, no. 4, p. 393-396. DOI: 10.1111/j.1502-3931.1970.tb00830.x
Seilacher A. (1973).- Fabricational noise in adaptive morphology.- Systematic Zoology, vol. 22, no. 4, p. 451-465.
Seilacher A. (1979).- Constructional morphology of sand dollars.- Paleobiology, vol. 5, no. 3, p. 191-221.
Seilacher A. (1989).- Vendozoa: Organismic construction in the Proterozoic biosphere.- Lethaia, Oslo, vol. 22, no. 3, p. 229-239. DOI: 10.1111/j.1502-3931.1989.tb01332.x
Seilacher, A. (1991).- Self-organizing mechanisms in morphogenesis and evolution. In: Schmidt-Kittler N. & Vogel K. (eds.), Constructional morphology and evolution.- Springer, Berlin, p. 251-271.
Seilacher A. (2007).- Trace fossil analysis.- Springer, Berlin, 226 p.
Shkolyar S., Bebout L., Blank J.G., Cady S.L, Cavalazzi B., Corbin E., Davila A.F., Des Marais D., Fisk M., Hickman-Lewis K., Lima-Zaloumis J., McLoughlin N., Murphy A.E., Noffke N., Perl S.M., Pohorille A., Potter-McIntyre S.L., Rainwater J.H. & Westall F. (2025).- Structural biosignatures - A category of potential biosignatures in the Life detection knowledge base.- Astrobiology, vol. 25, no. 7, p. 482-497.
Tenelanda-Osorio LI., Kappler A. & Mansor M. (2025).- Terrestrial iron biosignatures and their potential in Solar System exploration for astrobiology.- Earth-Science Reviews, vol. 271, article 105318, 21 p. DOI: 10.1016/j.earscirev.2025.105318
Thompson D'A.W. (1917/1942).- On growth and form.- Cambridge University Press, Cambridge - MA, 793 p. / 1116 p. URL: https://archive.org/details/ongrowthform00thom/page/770/mode/2up
Turing A.M. (1952).- The chemical basis of morphogenesis.- Philosophical Transactions of the Royal Society B, vol. 237, no. 641, p. 37-72. DOI: 10.1098/rstb.1952.0012
Vogel K. (1991).- Concepts of constructional morphology. In: Schmidt-Kittler N. & Vogel K. (eds.), Constructional morphology and evolution.- Springer, Berlin, p. 55-68.
Wagner G.P. (2014).- Homology, genes, and evolutionary innovation.- Princeton University Press, Princeton - NJ, 496 p.
Westall F., Foucher F., Bost N., Bertrand M., Loizeau D., Vago J.L., Kminek G., Gaboyer F., Campbell K.A., Bréhéret J.G., Gautret P. & Cockell C.S. (2015).- Biosignatures on Mars: What, where, and how? Implications for the search for Martian life.- Astrobiology, vol. 15, no. 11, p. 998-1029. DOI: 10.1089/ast.2015.1374
Westall F., Hickman-Lewis K., Cavalazzi B., Foucher F., Clodoré L. & Vago J.L. (2021).- On biosignatures for Mars.- International Journal of Astrobiology, vol. 20, no. 6, p. 377-393. DOI: 10.1017/S1473550421000264
Witten T.A. Jr & Sander L.M. (1981).- Diffusion-limited aggregation, a kinetic critical phenomenon.- Physical Review Letters, vol. 47, no. 19, p. 1400-1403.