◄ Carnets Geol. 26 (6) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Taxonomic notes]
[5. Biostratigraphic analysis]
[6. Definition of the Guadalupian Series of West Texas, USA]
[7. Previous biostratigraphic studies ... of the Guadalupian Series]
[8. Correlation of Olinal� Formation ammonoid assemblages ...]
[9. Conclusions]
[Bibliographic references] and ...
[Appendix]
Posgrado en Ciencias del Mar y Limnolog�a,
Universidad Nacional Aut�noma de M�xico, Ciudad Universitaria, 04510, Alcald�a
Coyoac�n, M�xico City (M�xico)
Escuela Superior de Ciencias de la
Tierra, Universidad Aut�noma de Guerrero, Ex-Hacienda de San Juan Bautista,
S/N, Taxco el Viejo, Guerrero (M�xico)
Instituto de Geociencias, Universidad
Nacional Aut�noma de M�xico, Quer�taro, Qro., 76230 (M�xico)
Departamento de Paleontolog�a, Instituto
de Geolog�a, Av. Universidad 3000, Circuito de la Investigaci�n Cient�fica,
Universidad Nacional Aut�noma de M�xico, 04510, Alcald�a Coyoac�n, Mexico
City (Mexico)
Departamento de Paleontolog�a, Instituto
de Geolog�a, Av. Universidad 3000, Circuito de la Investigaci�n Cient�fica,
Universidad Nacional Aut�noma de M�xico, 04510, Alcald�a Coyoac�n, Mexico
City (Mexico)
Published online in final form (pdf) on February 28, 2026
DOI 10.2110/carnets.2026.2606
![]()
[Editor:
Christina Ifrim; language editor: Robert W. Scott;
technical editor: Bruno R.C. Granier]
![]()
An ammonoid biostratigraphic analysis was conducted for the Permian Olinal� Formation, Guerrero state, southern Mexico. Bed-by-bed sampling yielded 113 ammonoids belonging to 14 different taxa. The biostratigraphic analysis allowed us to identify two ammonoid Zones: Paraceltites elegans and Coahuiloceras sp., and one subzone: Pseudagathiceras spinosum. According to the GSSP-defined boundaries, these biostratigraphic zones extend from the Roadian-Wordian to the lower Capitanian interval. To support the biostratigraphic analysis, several key taxa were reviewed from a taxonomic perspective. This taxonomic assessment validated the genus Coahuiloceras. A quantitative analysis based on morphometric ratios of the species of the genera Timorites and Coahuiloceras reveals a clear divergence between both groups. In addition, the biostratigraphic framework of the Olinal� succession is compared with previously published biostratigraphic data from Sonora, Coahuila and Texas, highlighting similarities and differences among these areas. The identified fauna in the Olinal� Formation is mainly associated with the American Realm.
� ammonoids;
� Middle Permian;
� Guadalupian;
� Olinal� Formation;
� biostratigraphy
Alanis-Pav�n A., Guerrero-Suastegui M., Ju�rez-Arriaga E., Torres-Mart�nez M.A. & Moreno-Bedmar J.A. (2026). Biostratigraphy of Guadalupian (Middle Permian) ammonoids from the Olinal� Formation, southern Mexico.- Carnets Geol., Madrid, vol. 26, no. 6, p. 125-150. DOI: 10.2110/carnets.2026.2606
Biostratigraphie des Ammono�d�s du Guadalupien (Permien moyen) de la Formation Olinal� (Mexique m�ridional).- Une analyse biostratigraphique fond�e sur les Ammono�d�s a �t� r�alis�e pour la Formation Olinal�, permienne, dans l'�tat de Guerrero (Mexique m�ridional). Un �chantillonnage banc par banc a permis de recueillir 113 sp�cimens appartenant � 14 taxons distincts. L'analyse biostratigraphique a permis d'identifier deux zones � Ammono�d�s, la Zone � Paraceltites elegans et et celle � Coahuiloceras sp., ainsi qu'une sous-zone, celle � Pseudagathiceras spinosum. D'apr�s les positions des limites d'�tages d�finies par les GSSP, ces zones biostratigraphiques s'�tendent de l'intervalle Roadien-Wordien jusqu'� la base du Capitanien. Afin d'�tayer l'analyse biostratigraphique, plusieurs taxons cl�s ont �t� r�examin�s du point de vue syst�matique. Cette r��valuation taxinomique a permis de valider le genre Coahuiloceras. Une analyse quantitative fond�e sur les rapports morphom�triques des esp�ces des genres Timorites et Coahuiloceras r�v�le une divergence nette entre les deux groupes. Par ailleurs, le cadre biostratigraphique de la succession � Olinal� est compar� aux donn�es biostratigraphiques pr�c�demment publi�es pour le Sonora, le Coahuila et le Texas, mettant en �vidence des similitudes et des diff�rences entre ces r�gions. La faune identifi�e dans la Formation Olinal� est principalement associ�e au domaine am�ricain.
� Ammono�d�s ;
� Permien moyen ;
� Guadalupien ;
� Formation Olinal� ;
� biostratigraphie
Through time, ammonoid biostratigraphy has been used as a reliable tool to establish broad and even global correlations among different geographic regions (Leonova, 2016). The first Paleozoic standard zonal scheme was based on ammonoids from the Permian Period, developed by B�se in 1919; but it was not until 1938 that Miller proposed an ammonoid biozonation for North America. Since then, several authors have refined these schemes depending on the distinct geographic regions (e.g., Ruzhencev, 1955; Glenister & Furnish, 1961; Jin et al., 1997; Bogoslovskaya et al., 1999; Leonova, 2011, 2016). This standard zonation has proven to be particularly useful for the Middle Permian in Mexico (Miller, 1944; Flores de Dios & Buitr�n-S�nchez, 1982; Villase�or-Mart�nez et al., 1987; Gonz�lez-Arreola et al., 1994; Alanis-Pav�n et al., 2024). Consequently, the first approach to define a Mexican Permian ammonoid zonation was made by Miller (1944) for the Paleozoic succession of the Las Delicias Formation, Coahuila, considered one of the most important regions with Permian cephalopods in Mexico. There, Miller (1944) detected the occurrence of the Perrinites Zone (Kungurian-Roadian), followed by the Waagenoceras Zone (Wordian), and Timorites Zone (Capitanian).
Nevertheless, the Las Delicias Formation is not the
only one where ammonoids occur in Mexico. Thus, we highlight the Olinal�
Formation exposed in northeastern Guerrero state, southern Mexico (Fig.
1 ![]() ),
whose Permian cephalopod fauna is abundant and diverse. Although the ammonoid
faunas of this unit have been considered worthy of attention worldwide, studies
on them are still scarce. In this context, only two works have focused on these
Permian ammonoids. The former was by Gonz�lez-Arreola
et al. (1994), who employed the invertebrates,
without providing systematic descriptions, to propose the Guadalupian for the
unit. Although the authors employed ammonoids to date the formation, the
inaccurate identification of the taxa only allowed them to relate the age of the
rocks to the Middle Permian. Later, Alanis-Pav�n
et al. (2024) reviewed and updated the taxonomy of
the ammonoids previously reported by Gonz�lez-Arreola
et al. (1994), rectifying the stratigraphic
position of particular ammonite-bearing rocks. Nonetheless, due to the poor
stratigraphic control of these ammonoids, it was not possible to establish more
reliable biostratigraphic ranges. It should be noted that Gonz�lez-Arreola
et al.
(1994) provided an initial framework for understanding the Olinal� Formation;
however, due to the limited availability of precise data, other methods were
employed to establish the stratigraphic position of the Olinal� Formation.
Thus, fusulinid biostratigraphy (Vachard et al.,
1993, 1997) and numerical dating using detrital zircon
U-Pb ages (Ju�rez-Arriaga & Murillo-Mu�et�n,
2020) were used, but these studies only included some specific strata.
),
whose Permian cephalopod fauna is abundant and diverse. Although the ammonoid
faunas of this unit have been considered worthy of attention worldwide, studies
on them are still scarce. In this context, only two works have focused on these
Permian ammonoids. The former was by Gonz�lez-Arreola
et al. (1994), who employed the invertebrates,
without providing systematic descriptions, to propose the Guadalupian for the
unit. Although the authors employed ammonoids to date the formation, the
inaccurate identification of the taxa only allowed them to relate the age of the
rocks to the Middle Permian. Later, Alanis-Pav�n
et al. (2024) reviewed and updated the taxonomy of
the ammonoids previously reported by Gonz�lez-Arreola
et al. (1994), rectifying the stratigraphic
position of particular ammonite-bearing rocks. Nonetheless, due to the poor
stratigraphic control of these ammonoids, it was not possible to establish more
reliable biostratigraphic ranges. It should be noted that Gonz�lez-Arreola
et al.
(1994) provided an initial framework for understanding the Olinal� Formation;
however, due to the limited availability of precise data, other methods were
employed to establish the stratigraphic position of the Olinal� Formation.
Thus, fusulinid biostratigraphy (Vachard et al.,
1993, 1997) and numerical dating using detrital zircon
U-Pb ages (Ju�rez-Arriaga & Murillo-Mu�et�n,
2020) were used, but these studies only included some specific strata.
Consequently, despite the effort to establish reliable ranges for the formation using other proxies, previous investigations did not result in precise correlations, only considering it as a Guadalupian unit. Hence, this work aims to study in detail the stratigraphy of the ammonoids recently collected in four different sections of the Olinal� Formation, making an accurate biostratigraphic analysis of the whole unit. This contribution enriches and complements the work started by Alanis-Pav�n et al. (2024), who resumed the study of Permian ammonoids of the Olinal� Formation, obtaining reliable relative temporal successions based on ammonoid assemblages. The establishment of an accurate stratigraphic position for the Olinal� Formation will enable the creation of reliable regional ammonoid zonation, allowing for the proposal of correlations with Coahuila and Texas assemblages, as well as considering a potential application for other Middle Permian formations where ammonoids from the American Realm are found.
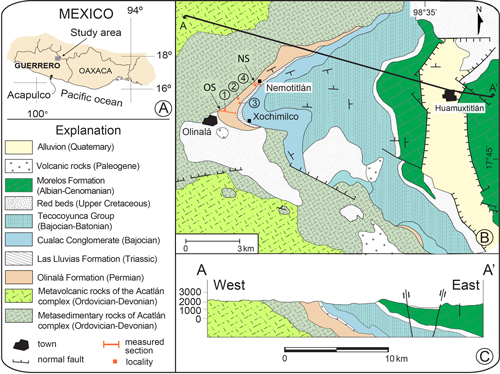
In this work, four stratigraphic sections from the
Olinal� Formation were studied (Fig. 1.B ![]() ). The first
section, Olinal�,
is located at 17�47'14.5"N 98�44'2.3"W, east of the town of
Olinal�, and is ~690 m thick. The second section, Terrerillos is located at 17�47'43.7"N 98�43'20.2"W, with a thickness of ~220 m. In both sections, the
stratigraphic record of the lower and middle parts of the formation are
observed. The third section, Barranca Coyomitla is located at 17�47'26.2"N 98�43'18.5"W, with a thickness of ~80 m. The fourth section, Nemotitl�n
is located at 17�48'13.2"N 98�42'43.3"W, 3 km northeast of the
town of Olinal�, displaying a thickness of ~860 m. The last two sections
represent the best outcrops with strata from the upper part of the formation.
). The first
section, Olinal�,
is located at 17�47'14.5"N 98�44'2.3"W, east of the town of
Olinal�, and is ~690 m thick. The second section, Terrerillos is located at 17�47'43.7"N 98�43'20.2"W, with a thickness of ~220 m. In both sections, the
stratigraphic record of the lower and middle parts of the formation are
observed. The third section, Barranca Coyomitla is located at 17�47'26.2"N 98�43'18.5"W, with a thickness of ~80 m. The fourth section, Nemotitl�n
is located at 17�48'13.2"N 98�42'43.3"W, 3 km northeast of the
town of Olinal�, displaying a thickness of ~860 m. The last two sections
represent the best outcrops with strata from the upper part of the formation.
|
Figure 1:
A) Geographic location of the study area, Guerrero state. B) Geologic map and studied sections. Geology
adapted from Campa et al. (1998); Juar�z-Arriaga
& Murillo-Mu�et�n (2020). C)
Structural cross-section A-A' (modified from Campa et al.,
1998); location of cross-section A-A' shown in
Fig. 1B. Sampling locations: A: Olinal�;
B: Terrerillos; C: Barranca Coyomitla; D: Nemotitl�n. |
Paleozoic outcrops of the Olinal� area were
reported for the first time in 1982, when Flores de Dios and Buitr�n-S�nchez
described the marine-origin clastic and carbonate sedimentary rocks in the
region, which were related to the Pennsylvanian-Permian. This range was
determined based on the fossil content and the stratigraphic position of the
sedimentary succession relative to the Lower Paleozoic metamorphic rocks of the
Acatl�n complex (Ortega-Guti�rrez et al., 1999), which constitutes the basement of the region
(Fig. 1.B-C ![]() ). Given that Flores de Dios and Buitr�n-S�nchez
(1982)
recognized the succession as a new unit made up of abundant limonitic
concretions in fine-grained marine clastic rocks, they suggested calling these
deposits the Olinal� Formation. Subsequently, Corona-Esquivel
(1983, 1985) restricted the formation range to the Upper Permian based on
identifying index fossils (e.g.,
ammonoids and brachiopods). In addition, he suggested that the Olinal�
Formation is overlain by the Las Lluvias Formation and the Cualac Conglomerate,
both of Jurassic.
). Given that Flores de Dios and Buitr�n-S�nchez
(1982)
recognized the succession as a new unit made up of abundant limonitic
concretions in fine-grained marine clastic rocks, they suggested calling these
deposits the Olinal� Formation. Subsequently, Corona-Esquivel
(1983, 1985) restricted the formation range to the Upper Permian based on
identifying index fossils (e.g.,
ammonoids and brachiopods). In addition, he suggested that the Olinal�
Formation is overlain by the Las Lluvias Formation and the Cualac Conglomerate,
both of Jurassic.
Corona-Esquivel (1983, 1985) subdivided the formation into seven lithologic units: 1) polymictic conglomerate of clasts, which are considered to be derived from the underlying the Acatl�n complex; 2) dark shale with intercalation of fine-grained sandstone, containing numerous concretions; 3) basal conglomerate and medium-to coarse-grained sandstone; 4) dark shale with limolitic concretions; 5) limestone; 6) alternating sandstone and shales beds with calcareous concretions; and 7) thick, fine-grained sandstone overlain by shale with concretions. It is worth mentioning that the Olinal� Formation has been mainly studied with an educational purpose for 30 years, through research projects and academic courses under the guidance of Mart�n Guerrero-Su�stegui from the Universidad Aut�noma de Guerrero. These projects have been focused on understanding and interpreting the depositional environments of the formation. After many years of hard work on the unit, the formation has been divided into four informal members: 1) conglomeratic member; 2) sandy member; 3) shaly and sandy shaly member; and 4) carbonate member (e.g., Guti�rrez-Quinto, 2002; Mor�n-Cuevas, 2007; Vega-Hern�ndez, 2011). Facies distribution of the Olinal� Formation indicates a complex sedimentary history that records a gradual transition from a siliciclastic coastal environment to a shallow carbonate platform (Ju�rez-Arriaga & Murillo-Mu�et�n, 2020). According to Ju�rez-Arriaga and Murillo-Mu�et�n (2020), the transitional change from shallow to deep marine deposits and the absence of reefs, slope deposits, and turbidites indicate that the Olinal� Formation was deposited on a homoclinal ramp developed in warm waters and normal salinity conditions.
The sedimentary succession comprising the
Olinal� Formation contains fossiliferous strata with a high diversity of fauna,
which have been reported in several studies and have contributed to establishing
the formation's stratigraphic range. Flores de Dios and Buitr�n-S�nchez
(1982) identified the brachiopods, Wellerella sp., Composita sp.,
and Paranorella sp., as
well as the ammonoids, Agathiceras freschi, Stacheoceras rothi, and Waagenoceras
sp., allowing a Late Carboniferous to Permian age to be
assigned to the fossiliferous strata. Likewise, the biota enabled the
establishment of a potential correlation with outcrops from the Mexican states
of Sonora, Hidalgo, and Chiapas. Later, Corona-Esquivel
(1983, 1985) made a more precise age-calibration of this formation, assigning a
Middle to Late Permian age based on the identification of ammonoids belonging to
the genera, Agathiceras, Paraceltites, Pseudogastrioceras, and
Stacheoceras, collected in limonitic concretions. He also
reported the presence of bivalves, gastropods, crinoids, and bryozoans.
Moreover, he proposed a correlation with coeval formations in Coahuila and
Chihuahua in Mexico. Subsequently, Vachard
et al. (1993, 2004) identified several microfossils,
including algae and small foraminifera, as well as the index fusulinids, Polydiexodina (erroneously interpreted as
Parafusulina bosei Dunbar
& Skinner, 1937, by Vachard
et al., 1993), Rauserella erratica Dunbar, 1944, and Codonofusiella extensa Skinner
& Wilde, 1955. These findings allowed Vachard
et al. (1993) to suggest that the base of the formation
corresponds to the Wordian Stage, whereas the upper stratigraphic part
corresponds to the Capitanian. Based on the proposals of Corona-Esquivel
(1983, 1985) and Vachard
et al. (1993), the Olinal� Formation was only
considered a Guadalupian unit. One year later, Gonz�lez-Arreola
et al. (1994) reported a faunistic assemblage
comprising the brachiopods, Thamnosia depressa, Cancrinella rugosa, Hustedia sp., and
Spiriferellina cristata, the bivalve Posidoniella sp., the conulariid
Paraconularia sp., the nautiloids Bitaunioceras coahuilensis Miller,
1944, and
the ammonoids Paraceltites
elegans Girty, 1908, Waagenoceras dieneri B�se,
1919, Pseudogastrioceras roadense (B�se,
1919), and Stacheoceras
toumanskyae Miller
& Furnish, 1940. The employment of this
assemblage and the identification of typical ammonoid species from the Middle
Permian enabled the authors to propose that the Olinal� Formation could be
correlated with the Las Delicias Formation in Coahuila, which is mainly
Guadalupian. Subsequently, Garibay-Romero et al.
(1998) reported
for the first time trilobite fragments in the formation, which material
consisted of an incomplete pygidium that they assigned to the species Anisopyge perannulata (Shumard,
1958), which belongs to the Cisuralian-Guadalupian. In 2000, Garibay-Romero
identified three ammonoid genera that had not been previously reported in the
formation: Doryceras Gemmellaro, 1887, Propinacoceras Gemmellaro,
1887, and Coahuiloceras Cant�-Chapa, 1997, all related to the Guadalupian Series. Likewise, Talavera-Mendoza et
al. (2005) provided
the first zircon dates from the Olinal� Formation, obtained from a quartz-rich
calcareous sandstone in the middle part of the type section; however, their
results indicated an Early Permian (Cisuralian) age. Later, Guerrero-S�nchez
and Quiroz-Barroso (2013) described bivalves from the shale facies, middle
part of the formation, confirming the previously proposed Middle Permian (Guadalupian)
range. Based on certain bivalve species identified, the depositional environment
was interpreted as a shallow, low-energy platform influenced by river sediment.
Recently, Ju�rez-Arriaga and Murillo-Mu�et�n
(2020) determined the maximum depositional age from U-Pb
detrital zircon analyses of sandstone samples (Nemo-2 and Nemo-4), obtained
near the base of the Olinal� Formation (Fig. 11 ![]() ). The maximum depositional
age for the oldest sandstone was established at 268�5 Ma (Roadian-Wordian),
coinciding with the stratigraphic range established from ammonoids previously
collected near the base of the unit. Finally, Alanis-Pav�n
et al. (2024) conducted a review and update of the material
reported by Gonz�lez-Arreola et al. (1994). In that work, the authors suggested that the
ammonoid faunal composition from the base to the middle part of the Olinal�
Formation was from the Wordian. Moreover, ammonoid specimens were assigned to
the American Realm, but these also indicated an affinity with coeval ammonoid
assemblages from the Paleotethys Realm.
). The maximum depositional
age for the oldest sandstone was established at 268�5 Ma (Roadian-Wordian),
coinciding with the stratigraphic range established from ammonoids previously
collected near the base of the unit. Finally, Alanis-Pav�n
et al. (2024) conducted a review and update of the material
reported by Gonz�lez-Arreola et al. (1994). In that work, the authors suggested that the
ammonoid faunal composition from the base to the middle part of the Olinal�
Formation was from the Wordian. Moreover, ammonoid specimens were assigned to
the American Realm, but these also indicated an affinity with coeval ammonoid
assemblages from the Paleotethys Realm.
The ammonoid sampling for this study was carried out in the September 2023 fieldwork campaign. Cephalopods were collected by standard sampling in black shale facies, ensuring meticulous biostratigraphic control. Ammonoids are generally preserved as calcareous molds in siltstone concretions. Due to the wet weather in the Olinal� area, Guerrero, conservation varies between complete and heavily fragmented shells. The generic and supra-generic-level taxonomy follows the proposals by Leonova (2002) and Furnish et al. (2009). Shell parameters were quantified using standard morphometric measurements, based on the approaches proposed by Korn (2010) and Klug et al. (2015), with certain modifications adapted for this study. The measured parameters include: Dm (shell diameter), wh (whorl height), ww (whorl width), uw (umbilical width), ww/wh (whorl width index), ww/Dm (shell width index), uw/Dm (umbilical width index), and wh/Dm (whorl height index) (Table 1). All dimensions are given in millimeters. Quantitative data were analyzed through the statistical freeware PAST version 4.07b (https://past.en.lo4d.com/windows). In addition, we measured and rephotographed some ammonoids collected by King (King et al., 1944) and described by Miller (1944), in particular those described as Timorites schucherti (Table 1). Ammonoids were coated with ammonium chloride prior to photographing. Additionally, we constructed one high-resolution 3D model using an EinScan-SP 3D scanner. The list of collected ammonoids, along with their identifications and IGM numbers, is provided in Appendix. All specimens were organized according to the fieldwork, including the bed number per section and specimen number (e.g., with the prefix: Oli1, Oli2, Te2, Nemo2, Nemo3, Coyo3). Types, figured, and other specimens examined in this study are deposited in the following institutions: Colecci�n Nacional de Paleontolog�a Mar�a del Carmen Perrilliat with the prefix IGM (Instituto Geol�gico Mexicano), Instituto de Geolog�a, Universidad Nacional Aut�noma de M�xico (UNAM), Mexico City, Mexico, and the Invertebrate Paleontology Collection at Yale Peabody Museum with the prefix YPM IP, New Haven, Connecticut, USA.
Table 1: Data parametrics of specimens of Timorites striatus, T. gemmellaroi, T. uddeni are taken from Miller and Furnish (1940) and Furnish and Glenister (1970). Data of T. schucherti (IGM 14328) are from Alanis-Pav�n et al. (2025). Note: Asterisk indicates approximate measurements.
| Species | Types | Dm | wh | ww | uw | wh/Dm | ww/Dm | uw/Dm | ww/wh |
| Timorites striatus | Delft 12754 | 21 | 6.93* | 10.5 | 11.5 | 0.33 | 0.5 | 0.55 | 1.51 |
| Timorites striatus | Iowa 12647 | 200 | 86* | 90* | 56* | 0.43 | 0.45 | 0.28 | 1.04 |
| Timorites striatus | Iowa 33572 | 135 | 64.8* | 64.8* | 36.4* | 0.48 | 0.48 | 0.27 | 1 |
| Timorites striatus | Iowa 31898 | 33 | 11.8 | 19.1* | 14.8* | 0.36 | 0.58 | 0.45 | 1.61 |
| Timorites gemmellaroi | No data | 162 | 64.8* | 128.5 | 26 | 0.4 | 0.79 | 0.16 | 1.98 |
| Timorites schucherti | YPM IP 16692 | 21.3 | 8.2 | 13.3 | 8.2 | 0.38 | 0.62 | 0.38 | 1.62 |
| Timorites schucherti | YPM IP 16693 | 29.5 | 10.7 | 16.3 | 9.7 | 0.36 | 0.55 | 0.32 | 1.52 |
| Timorites schucherti | YPM IP 16694 | 86.3 | 37.1 | 60.9 | 25.2 | 0.42 | 0.7 | 0.29 | 1.64 |
| Timorites schucherti | YPM IP 19133 | 34.1 | 10.8 | 21.5 | 12.8 | 0.31 | 0.63 | 0.37 | 1.99 |
| Timorites schucherti | IGM 14328 | 40.8 | 16.1 | 20.4* | 12.2 | 0.39 | 0.5 | 0.29 | 1.26 |
| Timorites uddeni | Iowa 1403 | 76 | 35.9* | 40 | 12 | 0.47 | 0.53 | 0.16 | 1.11 |
| Coahuiloceras sp. | IGM 14459 | 70.6 | 26.6 | 35.8 | 18.3* | 0.37 | 0.5 | 0.25 | 1.34 |
This section focuses on a brief discussion about biostratigraphically significance ammonoids with special attention on the taxa that were not previously taxonomically analyzed by Alanis-Pav�n et al. (2024). To support the taxonomic analysis, Appendix comprises a list of all studied ammonoids herein, including 120 specimens with the corresponding collection number, fieldwork sample name, and identifications, in addition to measurements of the more complete specimens.
The specimens assigned to Paraceltites elegans (Fig.
2.I-K
![]() ) are
well preserved, even better than those previously illustrated from the Olinal�
Formation (Gonz�lez-Arreola et al., 1994, Figs. c-e; Alanis-Pav�n
et al., 2024, Figs. 6.11-13, 10). A very conspicuous
characteristic of this genus is the evolute shell. This species exhibits a
rather robust, sinuous rib ornamentation on the evolute internal whorls. The
ribs have a low density and are strongest in the lower part of the flank. In the
last whorl, ribbing density abruptly increases, and the sinuous ribs become very
gracile. These traits allowed us to associate the material studied with typical
North American specimens of P. elegans (Miller & Furnish,
1940; King et al., 1944; Spinosa
et al., 1975; Alanis-Pav�n
et al., 2025).
) are
well preserved, even better than those previously illustrated from the Olinal�
Formation (Gonz�lez-Arreola et al., 1994, Figs. c-e; Alanis-Pav�n
et al., 2024, Figs. 6.11-13, 10). A very conspicuous
characteristic of this genus is the evolute shell. This species exhibits a
rather robust, sinuous rib ornamentation on the evolute internal whorls. The
ribs have a low density and are strongest in the lower part of the flank. In the
last whorl, ribbing density abruptly increases, and the sinuous ribs become very
gracile. These traits allowed us to associate the material studied with typical
North American specimens of P. elegans (Miller & Furnish,
1940; King et al., 1944; Spinosa
et al., 1975; Alanis-Pav�n
et al., 2025).
Adrianites sp. is represented by a single specimen, which is
poorly preserved and fragmented (Fig. 3.G ![]() ), but its main
ornamentation is still visible. The umbilicus is wide, evolute (uw/Dm= 0.52,
Table 2) with rounded shoulders. The phragmocone exhibits a
distinct, reticulate pattern, formed by longitudinal lirae and transverse lines,
characteristic of the genus Adrianites Gemmellaro,
1887. It is worth mentioning that the recrystallized
shell is broken in a part of the fragmocone that exposes the suture line (Fig.
3.G
), but its main
ornamentation is still visible. The umbilicus is wide, evolute (uw/Dm= 0.52,
Table 2) with rounded shoulders. The phragmocone exhibits a
distinct, reticulate pattern, formed by longitudinal lirae and transverse lines,
characteristic of the genus Adrianites Gemmellaro,
1887. It is worth mentioning that the recrystallized
shell is broken in a part of the fragmocone that exposes the suture line (Fig.
3.G ![]() ), which consists of a simple line with at least four
tongue-shaped lateral lobes and three rounded saddles. This suture line clearly
corresponds to the genus Adrianites (Fig. 4.A
), which consists of a simple line with at least four
tongue-shaped lateral lobes and three rounded saddles. This suture line clearly
corresponds to the genus Adrianites (Fig. 4.A ![]() ).
).
Table 2: Dimension (mm) and conch proportions of the most complete specimens. Ratios ww/wh: Whorl width index; ww/Dm: Shell width index; uw/Dm: Umbilical width index.
| Species | Types | wh/Dm | ww/Dm | uw/Dm | ww/wh |
| Paraceltites elegans | IGM 14443 | 0.25 | - | 0.52 | - |
| Adrianites sp. | IGM 14401 | 0.39 | - | 0.23 | - |
| Pseudagathiceras spinosum | IGM 14402 | 0.36 | 0.41 | 0.15 | 1.13 |
| Mexicoceras sp. | IGM 14541 | 0.31 | 0.94 | 0.27 | 3.02 |
Pseudagathiceras spinosum Miller,
1944 (Fig. 3.C-D ![]() ), was initially described in the Las Delicias
Formation, Coahuila, Mexico (King et al., 1944; Alanis-Pav�n
et al., 2025). Our specimens may represent a juvenile form,
exhibiting an involute shell with visible internal whorl constrictions (ww/Dm=
0.41; uw/Dm= 0.15, Table 2). The
ornamentation is characterized by longitudinal lirae, and the venter bears two
rows of spines. The suture is not visible. This report represents the second
documented occurrence of the species in a Mexican locality. Besides, it seems
that the spine density is a variable character because the specimen from
Coahuila (Miller, 1944, p. 103,
Pl. 29, figs.
1-5; Alanis-Pav�n et al.,
2025, p. 12, Fig. 7.4-7.7) has denser spines.
), was initially described in the Las Delicias
Formation, Coahuila, Mexico (King et al., 1944; Alanis-Pav�n
et al., 2025). Our specimens may represent a juvenile form,
exhibiting an involute shell with visible internal whorl constrictions (ww/Dm=
0.41; uw/Dm= 0.15, Table 2). The
ornamentation is characterized by longitudinal lirae, and the venter bears two
rows of spines. The suture is not visible. This report represents the second
documented occurrence of the species in a Mexican locality. Besides, it seems
that the spine density is a variable character because the specimen from
Coahuila (Miller, 1944, p. 103,
Pl. 29, figs.
1-5; Alanis-Pav�n et al.,
2025, p. 12, Fig. 7.4-7.7) has denser spines.
The material of Propinacoceras sp. (Fig.
3.A ![]() ) comprises a
single, very fragmented and small shell. Nonetheless, it is sufficiently
well-preserved to reveal the ornamentation. The venter displays a pair of
prominent, rounded node rows separated by a poorly developed median furrow.
These characteristics allowed us to make a generic assignment, but due to the
specimen preservation, obtaining a reliable specific identification was not
possible.
) comprises a
single, very fragmented and small shell. Nonetheless, it is sufficiently
well-preserved to reveal the ornamentation. The venter displays a pair of
prominent, rounded node rows separated by a poorly developed median furrow.
These characteristics allowed us to make a generic assignment, but due to the
specimen preservation, obtaining a reliable specific identification was not
possible.
|
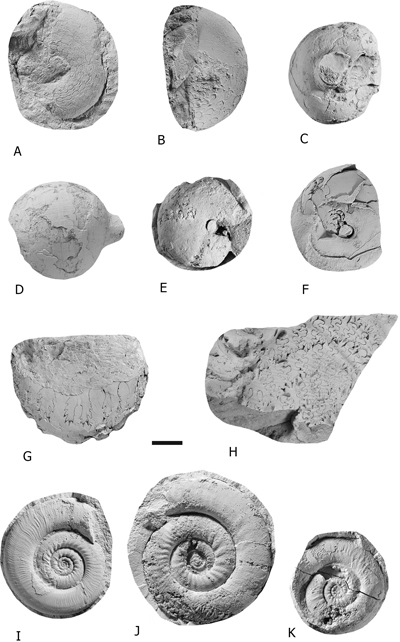
Figure 2:
A) Stacheoceras sp.,
lateral view, IGM 14497. B) Stacheoceras sp., lateral view, IGM 14489.
C-D)
Mexicoceras sp.,
lateral and ventral views, IGM 14451. E-G) Waagenoceras cf. W. dieneri (B�se,
1919),
lateral
and ventral views, IGM 14450, IGM 14481, IGM 14449. H) Waagenoceras cf. W. girtyi (B�se,
1919), lateral view, IGM 14471.
I-K)
Paraceltites elegans Girty, 1908, lateral views, IGM 14443, IGM 14409, IGM 14408. Scale bar = 1 cm. |
A single fragmentary specimen of Propinacoceras beyrichi Gemmellaro,
1887 (Fig. 3.B ![]() ), shows the flat flanks and part of the
venter.
The lateral view displays a very involute shell with a venter and a row of
rounded nodes. The suture consists of a bifid first-lateral lobe, while the
second lateral lobe is more elongated than the first. The lateral lobes are
small and bifid, and the saddles are rounded and slightly elongated (Fig.
4.B
), shows the flat flanks and part of the
venter.
The lateral view displays a very involute shell with a venter and a row of
rounded nodes. The suture consists of a bifid first-lateral lobe, while the
second lateral lobe is more elongated than the first. The lateral lobes are
small and bifid, and the saddles are rounded and slightly elongated (Fig.
4.B ![]() ). This taxon is described and illustrated for the
first time from the Guadalupian of Mexico. The traits mentioned correspond with
those recorded for the type species of the genus Propinacoceras. This taxon was described initially in the Wordian of
Sicily, Italy. Later, it was reported in other Wordian localities, including
British Columbia (Nassichuk et al., 1977) and South China (Zhou,
2017).
). This taxon is described and illustrated for the
first time from the Guadalupian of Mexico. The traits mentioned correspond with
those recorded for the type species of the genus Propinacoceras. This taxon was described initially in the Wordian of
Sicily, Italy. Later, it was reported in other Wordian localities, including
British Columbia (Nassichuk et al., 1977) and South China (Zhou,
2017).
The species, Waagenoceras cf. W. girtyi Miller
& Furnish, 1940 (Fig. 2.H ![]() ), is represented by two fragmentary specimens.
The inner molds show fine transverse lines. An incomplete suture is also
visible, highly arcuate, with at least seven lateral lobes. The lobes are
multiply digitated, and the saddles are mushroom-shaped (Fig.
4.E
), is represented by two fragmentary specimens.
The inner molds show fine transverse lines. An incomplete suture is also
visible, highly arcuate, with at least seven lateral lobes. The lobes are
multiply digitated, and the saddles are mushroom-shaped (Fig.
4.E ![]() ). Unfortunately, no complete specimens were found;
however, we were able to compare the suture of our samples with those
illustrated by Miller (1944, p. 113, Fig.
22C), Nassichuk (1977, p. 582, Fig. 17)
and Alanis-Pav�n et al.,
(2025, p. 21, Fig. 13.5, 17.3, 17.4, 18.1-18.4), concluding that
the Mexican specimens are associated with W. girtyi. Due to the preservation, it was not possible to make
a more precise assignment. This species was previously described in the
Wordian-Capitanian of Coahuila state and British Columbia.
). Unfortunately, no complete specimens were found;
however, we were able to compare the suture of our samples with those
illustrated by Miller (1944, p. 113, Fig.
22C), Nassichuk (1977, p. 582, Fig. 17)
and Alanis-Pav�n et al.,
(2025, p. 21, Fig. 13.5, 17.3, 17.4, 18.1-18.4), concluding that
the Mexican specimens are associated with W. girtyi. Due to the preservation, it was not possible to make
a more precise assignment. This species was previously described in the
Wordian-Capitanian of Coahuila state and British Columbia.
|
Figure 3:
A) Propinacoceras sp., ventral view, IGM 14439. B)
Propinacoceras beyrichi Gemellaro, 1887, lateral view, IGM 14468. C-D)
Pseudagathiceras spinosum Miller, 1944, lateral and ventral
views, IGM 14402. E-F) Agathiceras sp.,
lateral views, IGM 14426, IGM 14427. G) Adrianites sp.,
lateral view, IGM 14401. H) Epithalassoceras sp., lateral view, IGM 14440.
I-J)
Roadoceras roadense (B�se, 1919), lateral views, IGM 14404, IGM 14405. K) Metacrimites sp., lateral view, IGM 14441. Black triangles
emphasize two rows of spines at the beginning of the body chamber. Scale bar = 1
cm. |
Mexicoceras sp. (Fig. 2.C-D ![]() ) has
a strongly globular shell with a noticeably wide umbilicus (ww/Dm= 0.94; uw/Dm=0.27,
Table 2). The whorl section is depressed, with a
broad, rounded venter and low, strongly arched flanks. The ornamentation
consists of fine transverse lirae, visible in the preserved areas of the
recrystallized shell. Notably, some shell layers can be observed in some of
these recrystallized areas of the shell. The origin of these layers is difficult
to interpret, but at least three possible explanations can be considered: they
may result from taphonomic processes, diagenesis, or perhaps a preservation of
the original shell layering. The combination of a globular shell and fine
transverse lirae allows identifying the first occurrence of the genus Mexicoceras
Ruzhencev, 1955, in the Olinal� Formation. This genus
has been reported in the Guadalupian of the Delaware Basin (South Wells, Hegler,
and Pinery Limestone members) in Texas, USA (Miller & Furnish,
1940), as well
as in Wordian-Capitanian strata from Coahuila, Mexico (King et
al., 1944; Alanis-Pav�n
et al.,
2025).
) has
a strongly globular shell with a noticeably wide umbilicus (ww/Dm= 0.94; uw/Dm=0.27,
Table 2). The whorl section is depressed, with a
broad, rounded venter and low, strongly arched flanks. The ornamentation
consists of fine transverse lirae, visible in the preserved areas of the
recrystallized shell. Notably, some shell layers can be observed in some of
these recrystallized areas of the shell. The origin of these layers is difficult
to interpret, but at least three possible explanations can be considered: they
may result from taphonomic processes, diagenesis, or perhaps a preservation of
the original shell layering. The combination of a globular shell and fine
transverse lirae allows identifying the first occurrence of the genus Mexicoceras
Ruzhencev, 1955, in the Olinal� Formation. This genus
has been reported in the Guadalupian of the Delaware Basin (South Wells, Hegler,
and Pinery Limestone members) in Texas, USA (Miller & Furnish,
1940), as well
as in Wordian-Capitanian strata from Coahuila, Mexico (King et
al., 1944; Alanis-Pav�n
et al.,
2025).
Metacrimites sp. is represented by a poorly preserved
specimen (Fig. 3.K ![]() ), in which part of the external suture line is still visible.
The shell is involute, with a small, open umbilicus. The whorl section is
depressed, and the venter is rounded but slightly flattened ventrolaterally. The
lateral lobes are tongue-shaped and consist of at least five lobes that
gradually decrease in size toward the umbilicus. All saddles are rounded. Based
on its diagnostic features, this specimen could be a representative of the
Superfamily Adrianitinae. It should be noted that Cant�-Chapa
(1997) established the new genus Millerites, with a stratigraphic range that began in the Middle
Permian, distinguishing it from the previously used genus, Adrianites. Later,
Furnish et al. (2009)
regarded the genus Millerites as a
synonym of Neocrimites. However, Leonova and Boiko
(2015) recognized a lack of
revision in Ruzhencev's (1950) work on the
Superfamily Adrianitinae, highlighting that the diagnostic features were more
closely related to Metacrimites. Therefore, we assign the taxon analyzed herein
to the genus Metacrimites.
), in which part of the external suture line is still visible.
The shell is involute, with a small, open umbilicus. The whorl section is
depressed, and the venter is rounded but slightly flattened ventrolaterally. The
lateral lobes are tongue-shaped and consist of at least five lobes that
gradually decrease in size toward the umbilicus. All saddles are rounded. Based
on its diagnostic features, this specimen could be a representative of the
Superfamily Adrianitinae. It should be noted that Cant�-Chapa
(1997) established the new genus Millerites, with a stratigraphic range that began in the Middle
Permian, distinguishing it from the previously used genus, Adrianites. Later,
Furnish et al. (2009)
regarded the genus Millerites as a
synonym of Neocrimites. However, Leonova and Boiko
(2015) recognized a lack of
revision in Ruzhencev's (1950) work on the
Superfamily Adrianitinae, highlighting that the diagnostic features were more
closely related to Metacrimites. Therefore, we assign the taxon analyzed herein
to the genus Metacrimites.
|
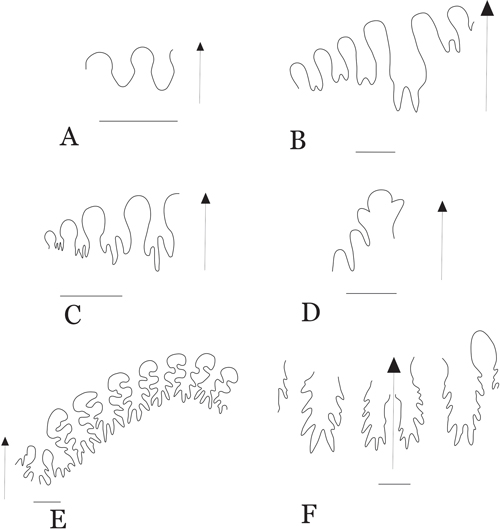
Figure 4:
External sutures: A) Adrianites sp., based on sample IGM
14401. B) Propinacoceras beyrichi, based on sample IGM 14468. C)
Stacheoceras sp.,
based on sample IGM 14489. D) Epithalassoceras sp., based
on sample IGM 14440. E) Waagenoceras cf. W. girtyi based on sample IGM 14471.
F)
Waagenoceras cf. W. dieneri, based on sample IGM 14449. Scale bar = 5 mm. |
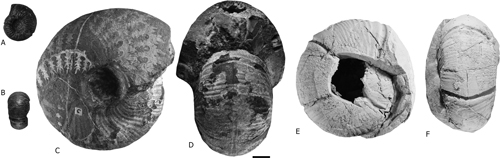
Coahuiloceras sp. (Fig. 5.E-F ![]() ) is reported for the first time in the Olinal�
Formation. This genus was proposed by Cant�-Chapa
(1997, p. 82, Fig. 46.E), who assigned it to the Subfamily Changhsingoceratinae.
In our opinion, the three studied specimens, herein, which include one well
preserved and complete specimen (Fig. 6
) is reported for the first time in the Olinal�
Formation. This genus was proposed by Cant�-Chapa
(1997, p. 82, Fig. 46.E), who assigned it to the Subfamily Changhsingoceratinae.
In our opinion, the three studied specimens, herein, which include one well
preserved and complete specimen (Fig. 6 ![]() ;
stl file download at https://carnetsgeol.net/cg/26/06/CG2606_Fig_06.stl
;
stl file download at https://carnetsgeol.net/cg/26/06/CG2606_Fig_06.stl ![]() ), must be only
classified in the Family Cyclolobidae. The thick discoidal shell (wh/Dm=0.37, uw/Dm=0.25),
with arcuate transverse striae and prominent sinuous constrictions are
diagnostic features of the Family Cyclolobidae. It is worth noting that Miller
(1944) illustrated a similar specimen (Fig. 5.C-D
), must be only
classified in the Family Cyclolobidae. The thick discoidal shell (wh/Dm=0.37, uw/Dm=0.25),
with arcuate transverse striae and prominent sinuous constrictions are
diagnostic features of the Family Cyclolobidae. It is worth noting that Miller
(1944) illustrated a similar specimen (Fig. 5.C-D ![]() ), but it was identified as
Timorites schucherti (Fig. 5.A-B
), but it was identified as
Timorites schucherti (Fig. 5.A-B ![]() ). In 2019, one of us re-examined and re-photographed
(A.A.P.)
the ammonoid collection originally sampled by King (King et al.,
1944) and later described by Miller
(1944). Therefore, we believe that this reconsideration is fully
supported due to the direct comparison with Miller's specimens. As shown in
Figure 5.C-F
). In 2019, one of us re-examined and re-photographed
(A.A.P.)
the ammonoid collection originally sampled by King (King et al.,
1944) and later described by Miller
(1944). Therefore, we believe that this reconsideration is fully
supported due to the direct comparison with Miller's specimens. As shown in
Figure 5.C-F ![]() , the specimens illustrated, herein, must be reassigned
to the genus Coahuiloceras.
Regarding species identification, further data are necessary to know the
specific content of this genus.
, the specimens illustrated, herein, must be reassigned
to the genus Coahuiloceras.
Regarding species identification, further data are necessary to know the
specific content of this genus.
|
Figure 5:
A-D) Timorites schucherti, lateral, apertural and ventral views, YPM IP
16692 and 16694. E-F) Coahuiloceras sp., lateral and ventral views, IGM 14459.
Scale bar = 1 cm. |
|
Figure 6:
Coahuiloceras sp.,
3D model, based on sample IGM 14459 (stl file download at https://carnetsgeol.net/cg/26/06/CG2606_Fig_06.stl |
Therefore, to support the qualitative
morphological difference between Timorites schucherti and Coahuiloceras sp.,
we consider a quantitative morphological analysis based on the following
morphometric ratios to be significant: wh/Dm, ww/Dm, uw/Dm, and ww/wh (Table
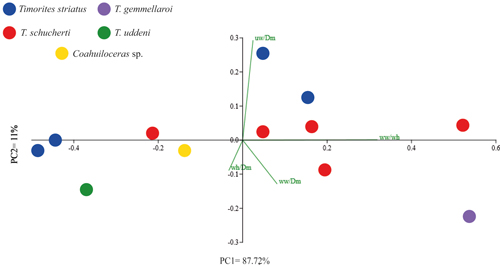
1). The first two Principal Components Analysis (PCA),
based on the whorl ratios width/whorl height (ww/wh) and umbilical
width/diameter (uw/Dm), explain 98.72% of the total variance (PC1 = 87.72%; PC2
= 11.00%;
Fig. 7 ![]() ). The PCA indicates that the variable with the
most significant influence in distinguishing the morphology of Coahuiloceras sp. and
Timorites schucherti is the ww/wh ratio, which may play a key role in their
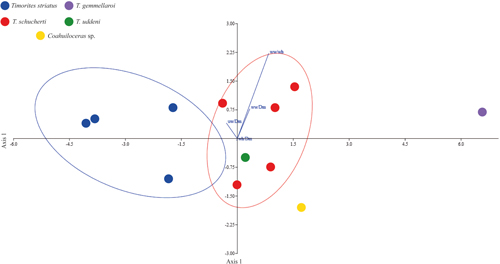
differentiation. To support previous results, we conducted a Linear Discriminant
Analysis (LDA), which revealed a clear morphological difference among the
analyzed species, particularly between Coahuiloceras sp. and most Timorites
species (Fig. 8
). The PCA indicates that the variable with the
most significant influence in distinguishing the morphology of Coahuiloceras sp. and
Timorites schucherti is the ww/wh ratio, which may play a key role in their
differentiation. To support previous results, we conducted a Linear Discriminant
Analysis (LDA), which revealed a clear morphological difference among the
analyzed species, particularly between Coahuiloceras sp. and most Timorites
species (Fig. 8 ![]() ). The first discriminant axis (LD1) explains
91.24% of the total variance among groups. In comparison, LD2 and LD3 account
for 4.92% and 3.80%, respectively. This distribution suggests that most
morphological differences among species can be effectively summarized along a
single dimension, supporting the visual separation observed between groups in
the morphospace. Timorites striatus
forms a well-defined cluster, indicating a distinctive morphology within the
group. In contrast, Timorites schucherti
occupies an intermediate position within the morphospace, suggesting that it
shares morphological characteristics with other species of the genus and could
represent a transitional form of the genus. Coahuiloceras sp. is markedly distinct from the other specimens
studied, reinforcing its assignment as a separate genus. This suggests a
morphology clearly differentiated from Timorites
forms. Overall, the results of the discriminant analysis not only support the
morphological validity of Coahuiloceras as a
distinct genus but also reveal considerable heterogeneity within Timorites. This pattern may have important taxonomic
implications, particularly regarding species delimitation within the group.
). The first discriminant axis (LD1) explains
91.24% of the total variance among groups. In comparison, LD2 and LD3 account
for 4.92% and 3.80%, respectively. This distribution suggests that most
morphological differences among species can be effectively summarized along a
single dimension, supporting the visual separation observed between groups in
the morphospace. Timorites striatus
forms a well-defined cluster, indicating a distinctive morphology within the
group. In contrast, Timorites schucherti
occupies an intermediate position within the morphospace, suggesting that it
shares morphological characteristics with other species of the genus and could
represent a transitional form of the genus. Coahuiloceras sp. is markedly distinct from the other specimens
studied, reinforcing its assignment as a separate genus. This suggests a
morphology clearly differentiated from Timorites
forms. Overall, the results of the discriminant analysis not only support the
morphological validity of Coahuiloceras as a
distinct genus but also reveal considerable heterogeneity within Timorites. This pattern may have important taxonomic
implications, particularly regarding species delimitation within the group.
|
Figure 7:
The plot dispersion from the Principal Component
Analysis (PCA) illustrates morphological differentiation between Coahuiloceras sp. and most
Timorites
species in the analyzed material. Based on the whorl width/whorl height (ww/wh)
and umbilical width/diameter (uw/Dm) ratios, the analysis explains 98.72% of the
total variance. |
|
Figure 8:
The plot dispersion from the Linear Discriminant
Analysis (LDA) illustrates morphological differentiation between Coahuiloceras sp. and most
Timorites species in the analyzed material. The first
discriminant axis (LD1) accounts for 91.24% of the total variance among groups,
while LD2 and LD3 explain 4.92% and 3.80%, respectively. |
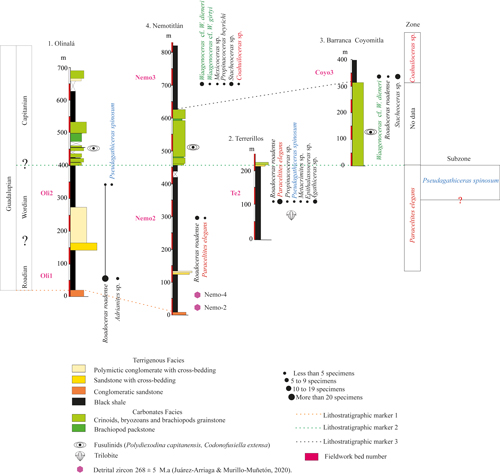
Ammonoid Zonation of the Olinal� Formation. The Guadalupian ammonoid assemblages of the
Olinal� Formation are mainly composed of previously described taxa from the
American Realm. Therefore, in this study, we considered the number of samples
per species collected bed by bed important, which enhanced the biostratigraphic
analysis. The use of interval biozones was preferred because the ammonoid record
of the Olinal� Formation allows reliable recognition of the lowest and highest
stratigraphic occurrences of key taxa (Fig. 9 ![]() ). This approach, consistent
with the definition of Murphy (1999) and Barrag�n et al.
(2010), provides objective criteria to establish zonal boundaries. According to
previous studies and the biostratigraphic data obtained in this work, we propose
the following markers for the Olinal� Formation:
). This approach, consistent
with the definition of Murphy (1999) and Barrag�n et al.
(2010), provides objective criteria to establish zonal boundaries. According to
previous studies and the biostratigraphic data obtained in this work, we propose
the following markers for the Olinal� Formation:
Lithostratigraphic marker 1: Ju�rez-Arriaga and Murillo-Mu�et�n (2020), based on U-Pb analyses of detrital zircon grains, providing a maximum depositional age of 268�5 Ma for the base of the sedimentary succession, wherewith the geochronological range may extend from the Roadian to the Capitanian;
Lithostratigraphic marker 2: Fusulinid biostratigraphy of carbonate facies, because the occurrence of Polydiexodina, Rauserella erratica, and Codonofusiella extensa in rudstone limestone indicates a minimum age of early to middle Capitanian for the fossil-bearing strata (Vachard et al., 1993, 1997, 2004);
Lithostratigraphic marker 3: Field observations indicating a transition from
carbonate to terrigenous facies (black shale with concretions) (Fig. 9 ![]() ).
Accordingly, we propose that the top of the carbonate interval denotes the base
of marker 3, further supported by ammonoid assemblages that allow reliable
correlation. These assemblages were grouped, in ascending order, into two zones
and one subzone, as follows.
).
Accordingly, we propose that the top of the carbonate interval denotes the base
of marker 3, further supported by ammonoid assemblages that allow reliable
correlation. These assemblages were grouped, in ascending order, into two zones
and one subzone, as follows.
|
Figure 9:
Biostratigraphical scheme of the Guadalupian with ammonoid distribution,
abundance and zonation of the Olinal� Formation. Taxa highlighted in red and
blue possess the most significant biostratigraphic value. Olinal� and Nemotitl�n
stratigraphic sections adapted from Juar�z-Arriaga and
Murillo-Mu�eton
(2020). |
Paraceltites elegans Zone. The index species is one of the most common ammonoids in the
lower and middle parts of the Olinal� Formation (Te2, Nemo2), below the shoal
marine platform limestone. The assemblage includes eight taxa: Paraceltites elegans,
Propinacoceras sp., Pseudagathiceras spinosum, Agathiceras sp.
(Fig. 3.E-F ![]() ),
Epithalassoceras sp. (Figs. 3.H
),
Epithalassoceras sp. (Figs. 3.H ![]() ,
4.D
,
4.D ![]() ), Roadoceras roadense (B�se,
1919)
(Fig. 3.I-J
), Roadoceras roadense (B�se,
1919)
(Fig. 3.I-J ![]() ),
Metacrimites sp. (Fig. 3.K
),
Metacrimites sp. (Fig. 3.K ![]() ), and Adrianites sp.
), and Adrianites sp.
Discussion: The specimens of Paraceltites elegans,
were collected in black shale from the middle part of the Terrerillos and Nemotitl�n sections
(Fig. 9.2-4 ![]() ). The oldest known representatives of the Order
Ceratitida, Family Paraceltitidae are attributable to the genus Paraceltites, whose first appearance (FA) occurred during
the Roadian (Leonova, 2002). Paraceltites elegans has been recorded in several Middle Permian
units of West Texas, with reported occurrences spanning the Capitanian. It
occurs in the Road Canyon Formation and the Williams Ranch Member of the Cutoff
Formation (Roadian), as well as in the Getaway Member (Lower Cherry Canyon
Formation) and the South Wells Limestone (Middle Cherry Canyon Formation) from
the Roadian-Wordian. Likewise, it has been found in the lower three members of
the Bell Canyon Formation (Hegler, Pinery, and Rader Limestone members), with a
stratigraphic distribution from the Wordian to the Capitanian. Its association
with diverse ammonoid faunas highlights its significance for Guadalupian
biostratigraphy. Likewise, we think Paraceltites elegans to be highly significant for the American Realm.
Besides, Paraceltites
elegans is phylogenetically
related to Cibolites
(Fig. 10.A
). The oldest known representatives of the Order
Ceratitida, Family Paraceltitidae are attributable to the genus Paraceltites, whose first appearance (FA) occurred during
the Roadian (Leonova, 2002). Paraceltites elegans has been recorded in several Middle Permian
units of West Texas, with reported occurrences spanning the Capitanian. It
occurs in the Road Canyon Formation and the Williams Ranch Member of the Cutoff
Formation (Roadian), as well as in the Getaway Member (Lower Cherry Canyon
Formation) and the South Wells Limestone (Middle Cherry Canyon Formation) from
the Roadian-Wordian. Likewise, it has been found in the lower three members of
the Bell Canyon Formation (Hegler, Pinery, and Rader Limestone members), with a
stratigraphic distribution from the Wordian to the Capitanian. Its association
with diverse ammonoid faunas highlights its significance for Guadalupian
biostratigraphy. Likewise, we think Paraceltites elegans to be highly significant for the American Realm.
Besides, Paraceltites
elegans is phylogenetically
related to Cibolites
(Fig. 10.A ![]() ), another index species used for the Capitanian
(Spinosa et al., 1975; Leonova, 2011).
), another index species used for the Capitanian
(Spinosa et al., 1975; Leonova, 2011).
This species has been associated with the following ammonoids: Agathiceras girtyi B�se, 1919, Epithalassoceras sp., Eumedlicotia burckhardti (B�se, 1919), Neocrimites sp., Perrinites vidriensis B�se, 1919, Propinacoceras sp., Stacheoceras sp., Texoceras texanum (Girty, 1908), Waagenoceras sp., Altudoceras altudense (B�se, 1919), Roadoceras beedei (Plummer & Scott, 1937), Roadoceras roadense (B�se, 1919), Mexicoceras guadalupense (Girty, 1908), Waagenoceras dieneri, Newellites richardsoni (Plummer & Scott, 1937), Cibolites uddeni Plummer & Scott, 1937, Timorites? sp., Neogeoceras girtyi (Miller & Furnish, 1940), Timorites cf. T. schucherti. (Miller & Furnish, 1940; Clifton, 1944; Spinosa et al., 1975; Lambert et al., 2000; Smith, 2012; Bell et al., 2015). In Mexico, King et al. (1944) recorded P. elegans in the Waagenoceras Zone and the Timorites Zone, corresponding to the Wordian and Capitanian, respectively.
Detrital zircon U-Pb ages reported constrain
the maximum depositional age at the base of the formation to 268�5 Ma (Roadian-Capitanian)
(Ju�rez-Arriaga & Murillo-Mu�et�n,
2020). This age range
coincides with the temporal framework proposed by Shen et
al. (2019)
for the earliest occurrences of Paraceltites in the Guadalupe Mountains.
Thus, the biostratigraphic framework established here for the Olinal� Formation
aligns with previously recognized ages, thereby reinforcing the reliability of
the P. elegans Zone as a useful proxy for constraining the
Roadian-Wordian boundary. The concordance between paleontological and
geochronological evidence underscores the potential of P. elegans to
serve as the basis for a regional zone with broader interregional correlation (Fig. 9 ![]() ).
).
|
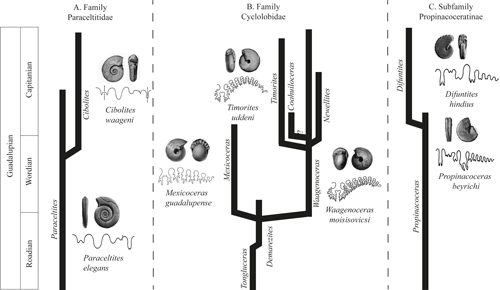
Figure 10:
Phylogenetic reconstruction of the A) Family Paraceltitidae, B)
Family Cyclolobidae, and C) Subfamily Propinacoceratinae. The arrows indicate that the lineages continue in the
stratigraphic record. Modified from Spinosa et al.
(1975) and Leonova
(2010). |
Coahuiloceras sp. Zone. This ammonoid zone is defined by first occurrence of
the index species. Coahuiloceras sp. occurs in the Nemotitl�n section, a taxon
previously considered a synonym of Timorites schucherti from Coahuila (see discussion above). This
genus was found in the upper part of the section, above the shoal marine
platform limestone (Nemo3 level). The ammonoid assemblage
includes Coahuiloceras sp., Waagenoceras cf. W. dieneri
(Figs. 2.E-G ![]() ,
4.F
,
4.F ![]() ), Waagenoceras cf. W. girtyi, Mexicoceras
sp. (Fig. 2.C-D
), Waagenoceras cf. W. girtyi, Mexicoceras
sp. (Fig. 2.C-D ![]() ), Propinacoceras beyrichi, Roadoceras
roadense, and Stacheoceras sp. (Fig. 2.A-B
), Propinacoceras beyrichi, Roadoceras
roadense, and Stacheoceras sp. (Fig. 2.A-B ![]() ).
Although Coahuiloceras sp. is only reported from the upper part of
the Nemotitl�n section (Nemo3), we infer that it may be coeval with the upper
part of the Barranca Coyomitla section (Coyo3), based on lithologic correlation
and the ammonoid assemblage found in this last section (e.g., Waagenoceras cf. W. dieneri, Stacheoceras sp.).
It should be noted that the thickness of strata in the two sections differs: in
Nemotitl�n, the black shale facies reach approximately 200 m, whereas in
Barranca Coyomitla, the black shale facies are less than 100 m thick.
Consequently, we suggest that Coahuiloceras sp.
was not found in the first 100 m of the Coyomitla section either, because its
first appearance lies above this interval, as observed in the Nemotitl�n
section.
).
Although Coahuiloceras sp. is only reported from the upper part of
the Nemotitl�n section (Nemo3), we infer that it may be coeval with the upper
part of the Barranca Coyomitla section (Coyo3), based on lithologic correlation
and the ammonoid assemblage found in this last section (e.g., Waagenoceras cf. W. dieneri, Stacheoceras sp.).
It should be noted that the thickness of strata in the two sections differs: in
Nemotitl�n, the black shale facies reach approximately 200 m, whereas in
Barranca Coyomitla, the black shale facies are less than 100 m thick.
Consequently, we suggest that Coahuiloceras sp.
was not found in the first 100 m of the Coyomitla section either, because its
first appearance lies above this interval, as observed in the Nemotitl�n
section.
Discussion: According to King et al. (1944), the complete faunal assemblage associated with Timorites schucherti (= Coahuiloceras sp.) from the Las Delicias Formation, Coahuila, included fusulinids (Polydiexodina capitanensis and Rauserella erratica) and ammonoids (Propinacoceras sp., Roadoceras roadense, Stacheoceras toumanskyae, Waagenoceras girtyi, Paraceltites elegans, and Cibolites uddeni).
The occurrence of fusulinids, such as Rauserella erratica, Codonofusiella extensa, and
Polydiexodina capitanensis, in the upper part of the Olinal� Formation suggests
a lower to middle Capitanian age for the strata (Vachard
et al., 1993, 1997,
2004), as these species are
typical late Guadalupian fusulinids of Texas. Polydiexodina capitanensis occurs in the Bell Canyon Formation (Pinery and Rader
Limestone members), while C. extensa is in
the McKittrick Canyon Limestone Member (Nestell
et al., 2019). These taxa are included in the Polydiexodina Zone, with a stratigraphic range from the
Wordian to the Capitanian, and in the Paraboultonia splendens Zone, which corresponds to the Capitanian,
respectively (Fig. 11 ![]() ) (Nestell et al.,
2019; Wahlman &
Nestell, 2024). The ammonoids in these members include Neogeoceras girtyi,
Mexicoceras guadalupense, and Paraceltites elegans (Miller & Furnish,
1940), supporting our record of the genus Mexicoceras in Capitanian strata.
) (Nestell et al.,
2019; Wahlman &
Nestell, 2024). The ammonoids in these members include Neogeoceras girtyi,
Mexicoceras guadalupense, and Paraceltites elegans (Miller & Furnish,
1940), supporting our record of the genus Mexicoceras in Capitanian strata.
|
Figure 11:
GSSPs
for the bases of the Roadian, Wordian, and Capitanian stages of the Guadalupian
Series in Texas. Each GSSP level coincides with the evolutionary appearance of
the conodont genus, Jinogondolella, and its correlation with fusulinid
and ammonoid biostratigraphy. ECM = El Centro Member; WRM = Williams Ranch
Member. Modified from Nestell et al. (2019), Henderson
and Shen (2020), and Wahlman
and Nestell (2024). |
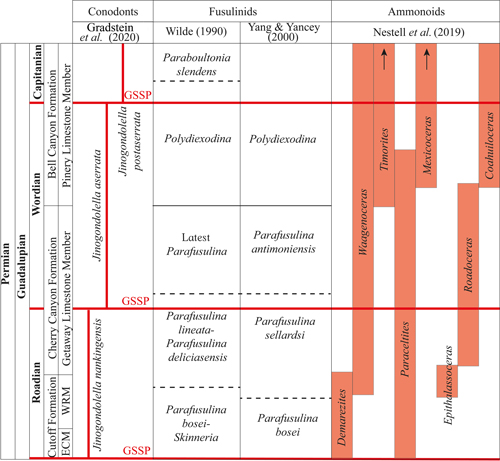
The occurrence of Waagenoceras cf. W. dieneri and Waagenoceras cf. W. girtyi allows extending the stratigraphic range of the genus into the early Capitanian, even though the genus has been considered an index component of the Wordian (Furnish, 1973; Jin et al., 1997; Leonova, 2011, 2018). This is supported by the previous record of Waagenoceras dieneri in the middle part of the Monos Formation in Sonora state, Mexico (Cooper et al., 1953), since at first, this ammonoid's presence suggested a Wordian range for the unit. However, a posterior analysis of the conodont assemblage composed of Jinogondolella nankingensis, J. aserrata, and J. postserrata enabled proposing an early to middle Capitanian range for the interval established as Wordian by Cooper et al. (1953; Lara-Pe�a et al., 2021).
Therefore, the presence of this genus would not
modify the proposed stratigraphic position, given that, according to Leonova
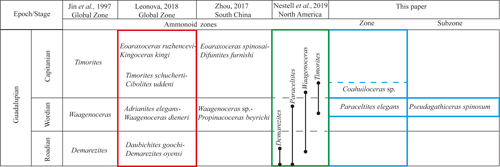
(2018), the lower boundary of the first Capitanian zone is defined by the Timorites schucherti-Cibolites uddeni
assemblage, while the upper boundary is marked by Eoaraxoceras ruzhencevi-Kingoceras kingi
(Fig. 12 ![]() ).
Similarly, Zhou (2017) established the upper
boundary based on the Eoaraxoceras spinosai-Difuntites furnishi assemblage
(Fig. 12
).
Similarly, Zhou (2017) established the upper
boundary based on the Eoaraxoceras spinosai-Difuntites furnishi assemblage
(Fig. 12 ![]() ).
).
In accordance with Leonova (2010, 2016), the lineage of the Family Cyclolobidae should be:
Tongluceras → Demarezites → Waagenoceras →
Cyclolobus → Changhsingoceras, with
the following iterations: Demarezites → Mexicoceras and Waagenoceras
→ Timorites and Newellites (Leonova, 2010, p.
269, Fig. 1). Consequently, we propose an adjustment to Leonova's iteration,
suggesting Demarezites → Mexicoceras and Waagenoceras
→ Timorites, Coahuiloceras, and Newellites. This proposal is founded on the genera
Timorites and Coahuiloceras, which are two partially
coeval offshoots of Waagenoceras.
Besides, the stratigraphic range of Coahuiloceras is clearly shorter of the two
(Fig. 10.B ![]() ), and it could be restricted to Mexico.
), and it could be restricted to Mexico.
A clear ancestor-descendant relationship may
exist between Coahuiloceras and Timorites,
as evidenced by the co-occurrence of Waagenoceras and Coahuiloceras.
However, Waagenoceras and Mexicoceras are typically assigned to the Wordian (Miller
& Furnish, 1940; Smith,
2012; Leonova, 2011, 2016). The proposed ancestor-descendant relationship is
based solely on the coexistence of Waagenoceras and Coahuiloceras. The
presence of species such as Waagenoceras cf. W. dieneri, Waagenoceras cf.
W. girtyi, and Mexicoceras sp. in strata of the Olinal� Formation could only
represent their last occurrences in North America, rather than evidence of an
evolutionary transition (Fig. 10.B ![]() ).
).
Pseudagathiceras spinosum Subzone. The base of this ammonoid subzone is defined by the first occurrence of taxa in the middle part of the Olinal� Formation, Terrerillos section, just below the shoal marine platform limestone beds. This subzone ranges from the middle to the upper part of the Paraceltites elegans Zone, coinciding with the ACME of P. elegans.
Discussion: We consider P. spinosum an endemic taxon, because previously it was described only from the Las Delicias Formation, Coahuila, within the Waagenoceras Zone (King et al., 1944). Recently, Alanis-Pav�n et al. (2025) described a juvenile specimen of this taxon corresponding to the Waagenoceras dieneri-Adrianites elegans Zone. Thus, the ammonoid assemblage is related to the Wordian Stage. The ammonoid assemblage associated with P. spinosum from Coahuila is similar to that found in the present study, including Roadoceras roadense, Epithalassoceras ruzencevi, Eumedlicottia burckhardti, Neogeoceras girtyi, Agathiceras girtyi, Stacheoceras gemmellaroi Miller, 1944, Mexicoceras guadalupense, M. smithi, Waagenoceras dieneri, W. girtyi, and Paraceltites elegans (King et al., 1944; Alanis-Pav�n et al., 2025).
The occurrence of P. spinosum in the P. elegans Zone, thus, provides higher resolution to the biostratigraphic framework of the Olinal� Formation. Its restricted stratigraphic distribution, which consistently co-occurs with a well-defined ammonoid assemblage and its recurrence across Coahuila and Olinal�, indicate that P. spinosum has considerable potential as a subzonal marker. Recognition of a P. spinosum Subzone would, therefore, improve Guadalupian ammonoid biostratigraphy in Mexico and provide a valuable tool for correlating sequences between the Olinal� and Las Delicias formations.
|
Figure 12:
Comparison of the ammonoid standard zones: global
zonation (Jin et al., 1997; Leonova,
2018), Tethys Realm (Zhou,
2017), American Realm (Nestell et al.,
2019), and the ammonoid zone of the Olinal� Formation. |
According to Grant et al. (2000), Hall in 1856 first discovered Permian
fossils in North America, which were initially related to the Carboniferous
Period. Later, the Shumard brothers (1858) collected fossils from the
Guadalupe Mountains in Texas, which were recognized as part of the Permian
fauna. Nonetheless, it was Girty (1902) who first proposed the term
Guadalupian, based on the Guadalupe Mountain outcrops. Subsequently, Girty
(1908) published a detailed account of the richness and diversity of the
Guadalupian fauna from the Guadalupe Mountains, revealing differences in faunal
composition between stratigraphic intervals. This background enabled the
establishment of the formal definition of the Middle Permian corresponding to
the Guadalupian Series, which comprises the Roadian, Wordian, and Capitanian
stages (Guadalupian Series) (Glenister et al., 1992,
1999; Grant
et al., 2000). The nominal reference formations of these stages are the
Road Canyon and Word formations in the Glass Mountains; similarly, the
Capitanian Stage is named after the massive Capitan Reef of the Guadalupe
Mountains, all located in Texas, USA (Glenister et al.,
1992). It
should be noted that the type area of this series is located in Guadalupe
Mountains National Park, where the rocks are well-exposed, mainly in West Texas
(Grant et al., 2000). In addition, the definition of the
Guadalupian Series was primarily based on the evolutionary sequence of a single
conodont genus: Jinogondolella (Glenister et al.,
1999;
Henderson & Shen, 2020) (Fig. 13 ![]() ).
).
The Roadian Stage is the basal stage of the Guadalupian Series and includes the El Centro Member of the Cutoff Formation (Glenister et al., 1999; Henderson & Shen, 2020). The marker horizon is the first appearance of the conodont Jinogondolella nankingensis (Henderson & Shen, 2020). The age of the base is estimated at 273.01 � 0.14 Ma based on U-Pb geochronological studies (Shen et al., 2020). The base of the Wordian Stage is identified in the upper part of the Getaway Limestone Member of the Cherry Canyon Formation (Glenister et al., 1999; Henderson & Shen, 2020). The marker horizon corresponds to the first appearance of the conodont Jinogondolella aserrata (Henderson & Shen, 2020). The age of the base is estimated at 266.9 � 0.4 Ma (Shen et al., 2020). The base of the Capitanian Stage is identified in the Pinery Limestone Member of the Bell Canyon Formation (Glenister et al., 1999; Henderson & Shen, 2020). The marker horizon corresponds to the first appearance of the conodont Jinogondolella postserrata (Henderson & Shen, 2020). The age of the base is estimated at 264.28 � 0.16 Ma (Shen et al., 2020).
The Guadalupian Series is divided into three
stages: Roadian, Wordian and Capitanian, which are defined by the first
appearances of different index conodonts. Nevertheless, other index fossils have
been used worldwide to correlate distinct outcrops associated with the
Guadalupian Series (Lambert et al., 2002; Leonova,
2009; Wu
et al., 2017; Chen & Xu,
2019; Shen et al., 2019). The use of other fossil groups has presented difficulties for
biostratigraphic correlations, mainly due to biogeographic provincialism or the
lack of data in the fossil record (Nestell et al.,
2019). For
example, although fusulinids are excellent index fossils, these are
geographically restricted, making comparisons between North American basins and
the Tethys region challenging (Huang et al., 2017; Zhang
& Wang, 2018; Nestell et al.,
2019; Arefifard
& Clapham, 2021; Wahlman & Nestell,
2024). Most
Guadalupian North American fusulinid data come from the Permian Basin of West
Texas (Wahlman & Nestell, 2024). The reported stratigraphic
ranges in early and middle Guadalupian fusulinids are somewhat inconsistent,
likely resulting from the limited amount of available data and from the
redefinition of Guadalupian Stage boundaries based on conodont biostratigraphy (Nestell
et al., 2019; Wahlman & Nestell,
2024)
(Fig. 11 ![]() ).
).
Regarding ammonoids, the challenges are
different, because Guadalupian ammonoid biostratigraphy has not been as well
established globally as that of conodonts or even fusulinids (Leonova,
2011; Shen et al., 2019,
2020). In the early 20th century, a
general Permian correlation for West Texas was established based on distinctive
ammonoid genera (Miller, 1938). In this sense, the ammonoid faunas of the
Roadian Stage were characterized by the appearance of Demarezites, the ancestral representative of the Cyclolobaceae,
which marked the onset of diversification of this group throughout the remainder
of the Guadalupian (Glenister et al., 1992,
1999). Likewise, Paraceltites
elegans
is present in most faunas, being dominant in West Texas, whereby its occurrence
near the base of this stage represents the appearance and initial
diversification of the Ceratitida (Glenister et al.,
1999; Shen
et al., 2019, 2020). Traditionally, the base of the Wordian Stage was
recognized in the Pipeline Shale Member at the base of the Brushy Canyon
Formation and was defined by the first occurrence of the genus Waagenoceras (Glenister et al.,
1992, 1999; Jin
et al., 1997). However, recent studies have shown that the ammonoid Waagenoceras also occurs in the upper Roadian Stage,
according to the current GSSP definitions (Shen et al.,
2019, 2020; Henderson & Shen,
2020) (Fig. 13 ![]() ). Similarly, the
base of the Capitanian Stage was traditionally recognized in the Hegler
Limestone Member of the Bell Canyon Formation and was characterized by the
occurrence of the genus Timorites (Glenister
et al., 1992, 1999; Jin et al.,
1997), although this genus
has also been collected in the middle Guadalupian Manzanita Member of the Cherry
Canyon Formation (Glenister et al., 1999). Thus, recent studies
have proposed that Timorites occurs from the
upper Wordian, as the currently defined GSSP (Nestell et al.,
2019; Shen et al., 2019,
2020; Henderson & Shen,
2020) (Fig. 13
). Similarly, the
base of the Capitanian Stage was traditionally recognized in the Hegler
Limestone Member of the Bell Canyon Formation and was characterized by the
occurrence of the genus Timorites (Glenister
et al., 1992, 1999; Jin et al.,
1997), although this genus
has also been collected in the middle Guadalupian Manzanita Member of the Cherry
Canyon Formation (Glenister et al., 1999). Thus, recent studies
have proposed that Timorites occurs from the
upper Wordian, as the currently defined GSSP (Nestell et al.,
2019; Shen et al., 2019,
2020; Henderson & Shen,
2020) (Fig. 13 ![]() ).
).
Various authors have employed this ammonoid zonation for global biostratigraphy. However, in recent years, the knowledge about these index fossils has been expanded by the inclusion of new data and review of previous information, including new taxonomic features and recognition of variations in stratigraphic and paleobiogeographic distribution of the faunal assemblages (Leonova, 1999, 2002, 2009, 2011, 2016, 2017, 2018; Ehiro & Misaki, 2005; Tazawa et al., 2005; Ehiro, 2008, 2022; Zhou, 2017; Alanis-Pav�n et al., 2025). Consequently, Ehiro and Araki (1997), Ehiro and Misaki (2005), and Ehiro (2008, 2022) identified Guadalupian ammonoid faunas in the Paleotethys Realm of northeastern Japan, describing assemblages whose genera, Waagenoceras and Timorites, allow the correlation of strata with typical regions of the Wordian and Capitanian, respectively. But Zhou (2017) noticed that ammonoids of the Paleotethys Realm from southern China were commonly endemic, making it difficult to compare the faunas studied with those found in other paleogeographic areas. Zhou (2017) also explained that endemic ammonoid assemblages were found on the southern China platform, noting that more cosmopolitan faunas were on the slope of the open basin areas. It is worth noting that Leonova (2011) recognized the difficulty in developing a Permian ammonoid zonal scale, because ammonoid localities are rare, and the transitions between species and genera that characterize long geological intervals are uncommon. According to Leonova (2011), a Permian ammonoid zonal stratigraphy is neither established nor accepted. Nevertheless, Leonova proposed a zonal subdivision based on a short stratigraphic interval, in a broad geographic range, and an evolutionary development that allows for comparing species of the same genus (Leonova, 2011). This proposal uses two isochronous index species across different biogeographic regions (Leonova, 2018). On the contrary, based on the open marine ammonoid assemblages, Zhou (2017) proposed one zonation for the Guadalupian, which enables comparing different regions worldwide: the Waagenoceras sp.-Propinacoceras beyrichi Zone for the Wordian, and the Eoaraxoceras spinosai-Difuntites furnishi Zone for the Capitanian.
All fusulinid biostratigraphic data from
Permian Basin formations in Texas, USA (Nestell et al.,
2019),
together with the species described by Vachard et al.
(1993, 2004)
from Olinal�, and the currently accepted conodont-based stages, are used in the
preceding discussion. These datasets can be approximately correlated with the
stratigraphic ranges of ammonoids shown in Figure 11 ![]() .
.
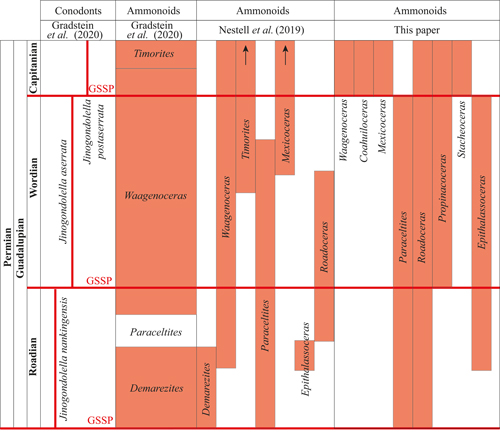
These contrasting biostratigraphic approaches highlight the need for new regional data, such as those presented in this study, which provide additional evidence to refine the Guadalupian ammonoid zonation for North America. The discrepancies between the classic ammonoid zones and the current GSSP framework indicate that the newly recognized stratigraphic ranges of Waagenoceras and Timorites require a reassessment of previous correlations.
The new Mexican ammonoid biostratigraphic data
generated in this study are highly consistent with that of Nestell et
al. (2019). The new Mexican data clearly demonstrate that the current
Wordian/Capitanian GSSP and its correlation with ammonoid zones can be refined.
In particular, the stratigraphic ranges of the genera Waagenoceras and Timorites
appear to be incorrectly represented in the current version of the Geological
Time Scale (GTS 2020; updated online by ICS, 2026 version). Here, we combine the Mexican
biostratigraphic data from this study with the North American data of Nestell
et al. (2019) (Fig. 13 ![]() ). Some inconsistency observed between these
datasets concerns the range of Paraceltites, which is extended slightly
higher in Mexico, with its last occurrence very close to the Wordian/Capitanian
GSSP. Based on the coherent data and this minor discrepancy in the range of Paraceltites,
we propose a new global ammonoid zonation that integrates the Roadian/Wordian
and Wordian/Capitanian GSSPs (Fig. 13
). Some inconsistency observed between these
datasets concerns the range of Paraceltites, which is extended slightly
higher in Mexico, with its last occurrence very close to the Wordian/Capitanian
GSSP. Based on the coherent data and this minor discrepancy in the range of Paraceltites,
we propose a new global ammonoid zonation that integrates the Roadian/Wordian
and Wordian/Capitanian GSSPs (Fig. 13 ![]() ). This new proposal represents a significant step
forward in refining the global biostratigraphic calibration of Middle Permian
ammonoids and enhances their potential for accurately dating Middle Permian
sedimentary successions.
). This new proposal represents a significant step
forward in refining the global biostratigraphic calibration of Middle Permian
ammonoids and enhances their potential for accurately dating Middle Permian
sedimentary successions.
The global Guadalupian ammonoid zonation proposed by Leonova
(2011, 2018) provides a refined framework that incorporates the zonation of Jin
et al. (1997), employing index species with broader geographic
distributions. A key improvement in this scheme is the subdivision into lower
and upper parts of the Capitanian Stage based on ammonoid assemblages, which
enhances biostratigraphic resolution. However, inconsistencies remain in the
stratigraphic ranges of Guadalupian ammonoid species, mainly due to the conodont-based
redefinition of stage boundaries (Henderson & Shen,
2020).
Being so, the present work aims to correlate the Guadalupian stages using
ammonoid and fusulinid-based chronostratigraphic units in comparison with the
currently accepted conodont-based framework (Fig. 11 ![]() ). The Middle Permian
strata, defined initially and exposed in the Guadalupe Mountains area, can be
recognized globally through the succession of conodonts, fusulinids, and
ammonoids. Nevertheless, the revised conodont-based Guadalupian boundaries
differ from the traditional ammonoid-based scheme, resulting in some correlation
inconsistencies within the Permian Basin and its equivalent regions (Fig. 11
). The Middle Permian
strata, defined initially and exposed in the Guadalupe Mountains area, can be
recognized globally through the succession of conodonts, fusulinids, and
ammonoids. Nevertheless, the revised conodont-based Guadalupian boundaries
differ from the traditional ammonoid-based scheme, resulting in some correlation
inconsistencies within the Permian Basin and its equivalent regions (Fig. 11 ![]() ).
).
On the other hand, Zhou's ammonoid zonation from the Nanpanjiang Basin, South China,
incorporates Wordian and upper Capitanian data. Zone 5 (Waagenoceras sp.-Propinacoceras beyrichi) corresponds to the
Wordian, while Zone 6 (Eoaraxoceras spinosai-Difuntites
furnishi) is assigned to the upper
Capitanian (Fig. 12 ![]() ) (Zhou,
2017). A
key point to consider is that Leonova (2002)
recognized the lineage Propinacoceras → Difuntites.
According to this data, during the Wordian, Propinacoceras beyrichi experienced a radiation event, giving rise to the
genus Difuntites Glenister
& Furnish, 1988, within the Subfamily Propinacoceratinae, marking the last known occurrence of this subfamily
(Fig. 10.C
) (Zhou,
2017). A
key point to consider is that Leonova (2002)
recognized the lineage Propinacoceras → Difuntites.
According to this data, during the Wordian, Propinacoceras beyrichi experienced a radiation event, giving rise to the
genus Difuntites Glenister
& Furnish, 1988, within the Subfamily Propinacoceratinae, marking the last known occurrence of this subfamily
(Fig. 10.C ![]() ). The genus
Difuntites has been identified in Capitanian strata (Ruzhencev,
1976; Cant�-Chapa, 1997). This is
particularly significant given that the Mexican species, P. beyrichi, has been reported from the lower
Capitanian, possibly
representing its last occurrences, while in South China, Difuntites furnishi (Zhou,
2017)
has been recorded from the upper Capitanian.
). The genus
Difuntites has been identified in Capitanian strata (Ruzhencev,
1976; Cant�-Chapa, 1997). This is
particularly significant given that the Mexican species, P. beyrichi, has been reported from the lower
Capitanian, possibly
representing its last occurrences, while in South China, Difuntites furnishi (Zhou,
2017)
has been recorded from the upper Capitanian.
Therefore, we propose that the biostratigraphic record of the taxa, Waagenoceras cf. W. dieneri, Waagenoceras cf. W. girtyi, Mexicoceras sp., and Propinacoceras beyrichi, should extend at least into the lower Capitanian, possibly representing their last occurrences.
In contrast, the Mexican ammonoid zonation
cannot be correlated with Leonova's and Zhou's scheme, because the
stratigraphic ranges of several species contradict our data. This discrepancy is
particularly evident in the Wordian, since the Mexican species Waagenoceras cf.
W. dieneri, Waagenoceras cf. W. girtyi, and Propinacoceras beyrichi have also been found in lower Capitanian strata.
Similarly, Alanis-Pav�n et al. (2025) reported Waagenoceras as ranging throughout the
ammonoid-based
Wordian and Capitanian stages. In contrast, Nestell et al.
(2019)
documented Waagenoceras ranging through the Roadian and Wordian
stages, and Timorites through the Wordian and Capitanian stages,
according to the conodont-based framework (Fig. 13 ![]() ).
).
|
Figure 13:
Guadalupian Series time scale showing GSSP conodont
markers compared to the ammonoid standard biostratigraphy, modified from Nestell
et al. (2019) and Gradstein et al. (2020). |
According to our biostratigraphic data, the Olinal� Formation can be correlated with Guadalupian formations of the American Realm. This statement supports previous studies on the Las Delicias Formation in Coahuila, the Cherry Canyon and Bell Canyon formations in Texas, USA, and the Cache Creek Group in British Columbia. Based on the studied ammonoid assemblages from the Olinal� Formation, the faunal composition and stratigraphic distribution suggest close affinities with the Guadalupian from West Texas.
It is also important to note the affinity between ammonoid assemblages from the American Realm and those from coeval units of South China and Japan. However, differences in taxonomic diversity between these two realms should also be noted. Such disparities may be attributed to paleobiogeographic and paleoecological factors, which warrant further investigation.
In the present work, two ammonoid zones and one subzone are recognized in the Olinal� Formation in the Guerrero state: the Paraceltites elegans Zone, Coahuiloceras sp. Zone and Pseudagathiceras spinosum Subzone. Several global ammonoid zonations for the Guadalupian have been proposed by different authors, many using Waagenoceras as a key index fossil for the Wordian and Timorites for the Capitanian. However, our finding of Waagenoceras in the upper levels of the Olinal� Formation (Guerrero, Mexico), which, based on fusulinids and the conodont-based framework, is assigned to the late Wordian-to early Capitanian, suggests that these biostratigraphic schemes may require reconsideration or refinement when applied to this region.
The biostratigraphic age proposed for the base of the formation, based on the P. elegans Zone, is consistent with the previously estimated geochronological range, Roadian-Wordian (268�5 Ma). Hence, we propose extending the biostratigraphic range of Waagenoceras cf. W. dieneri, Waagenoceras cf. W. girtyi, Mexicoceras sp., and Propinacoceras beyrichi at least into the lower Capitanian in the North American region (American Realm).
The taxon, Coahuiloceras sp., is validated, along with its phylogenetic relationship to Waagenoceras and Timorites. The quantitative analysis reinforces the recognition of Coahuiloceras as a morphologically distinct genus, while highlighting substantial morphological variability within Timorites. This observed pattern could carry significant taxonomic implications, particularly for defining species boundaries within the genus, because it can be assigned to the upper Wordian-lower Capitanian.
The ammonoids from the Olinal� Formation are assigned to the American Realm and exhibit faunal affinities with the Guadalupian ammonoid assemblages of the Guadalupe Mountains in West Texas and the Las Delicias Formation in Coahuila state, supporting regional biostratigraphic correlations. The integration of the new Mexican ammonoid data with that of Nestell et al. (2019) allows for a refined correlation of Permian ammonoid genera across the Roadian-Wordian and Wordian-Capitanian boundaries. This new proposal represents a significant step forward in improving the global biostratigraphic calibration of Middle Permian ammonoids and enhances their potential for accurately dating Middle Permian successions.
Alberto Alanis Pav�n thanks Posgrado de Ciencias del Mar y Limnolog�a of the Universidad Nacional Aut�noma de M�xico (UNAM) and the financial support provided by CONAHCYT through the scholarship number 931193 and gratefully acknowledges the members of his Ph.D. committee - M.A. Torres-Mart�nez, J.A. Moreno-Bedmar, R.J. Alc�ntara-Hern�ndez, P. Navas-Parejo, and J.F. S�nchez-Beristain - for their guidance and support. This article fulfills a requirement for the Doctorado en Ciencias del Mar y Limnolog�a, UNAM with a specialization in Geolog�a Marina, pursued by Alanis-Pav�n. MATM thanks the financial support supplied by the project PAPIIT-DGAPA-UNAM IN114623. We also extend our gratitude to the Division of Invertebrate Paleontology, Yale Peabody Museum because AAP received support from the Dunbar Grants in Aid Program for Invertebrate Paleontological Research. We also thank M.A. Argaez Mart�nez and D.A. Beltr�n L�pez for their assistance in preparing and photographing specimens. We would like to express our gratitude to Tatiana Leonova and an anonymous referee for their suggestions and comments, which greatly improved the original manuscript. We are particularly grateful to the Editor-in-Chief, Bruno Granier, for his support and guidance during the editorial process, and Christina Ifrim, for her time. The Senior Editor, Robert W. Scott, for his excellent editorial work. We especially thank him for the final handling of the manuscript and for his careful and thorough reading, which greatly contributed to improving the quality and clarity of the work.
Alanis-Pav�n A., Moreno-Bedmar J.A. & Torres-Mart�nez M.A. (2024).- Taxonomic and paleobiogeographic review of the Guadalupian ammonoids reported by Gonz�lez-Arreola et al. (1994) from the Olinal� Formation, Guerrero state, Mexico.- Journal of South American Earth Sciences, vol. 146, article 105072, 11 p.
Alanis-Pav�n A., Torres-Mart�nez M.A., Moreno-Bedmar J.A. & Utrup J. (2025).- Permian ammonoids from the Guadalupian (Wordian-Capitanian) of Las Delicias, Coahuila state, Mexico: New biostratigraphical and paleobiogeographical insights.- Journal of Paleontology, Cambridge (UK), vol. 99, no. 4, p. 857-889. DOI: 10.1017/jpa.2025.10140
Arefifard S. & Clapham M.E. (2021).- Cisuralian and Guadalupian global paleobiogeography of fusulinids in response to tectonics, ocean circulation and climate change.- Palæogeography, Palæoclimatology, Palæoecology, vol. 561, article 110052, 15 p.
Barrag�n R., Campos-Madrigal E., Ferrusqu�a-Villafranca I. & L�pez-Palomino I. (2010).- C�digo estratigr�fico norteamericano.- Bolet�n del Instituto de Geolog�a, Ciudad de M�xico, no. 117, p. 48 p. DOI: 10.22201/10.22201/igl.01855530e.2010.117
Bell G.L., Hearst J.M., Nestell M.K., Nestell G.P. & Lambert L.L. (2015).- Stratigraphic framework and biostratigraphic significance of the terminal Guadalupian reef Trail Member, Bell Canyon Formation in the Patterson hills, type Guadalupian area, Texas.- Stratigraphy, Flushing - NY, vol. 12, no. 2, p. 93-107.
Bogoslovskaya M.F., Kusina L.F. & Leonova T.B. (1999).- Classification and distribution of Late Paleozoic ammonoids. In: Rozanov A.Y. & Shevyrev A.A. (eds.), Fossil Cephalopods: Recent advances in their study.- Paleontological Institute, Russian Academy of Sciences, Moscow, p. 89-124 [in Russian].
B�se E. (1919).- The Permo-Carboniferous ammonoids of the Glass Mountains, West Texas, and their stratigraphical significance.- University of Texas Bulletin, Austin - TX, no. 1762, 241 p. URI: http://hdl.handle.net/2152/53803
Campa M.F., Garc�a-D�az J.L., Bustamante-Garc�a J., Torreblanca-Castro T.J., Aguilera-Mart�nez M.A. & Vergara-Mart�nez A. (1998).- Carta Geol�gico-Minera Chilpancingo E14-8 escala 1:250,000 estados de Guerrero, Oaxaca y Puebla: Pachuca, Hidalgo.- Servicio Geol�gico Mexicano, Aguascalientes, 1 map.
Cant�-Chapa A. (1997).- Los cefal�podos del Paleozoico de M�xico.- Geociencias, Ciudad de M�xico, no. 1, 127 p. (XV Pls.).
Chen J. & Xu Y.G. (2019).- Establishing the link between Permian volcanism and biodiversity changes: Insights from geochemical proxies.- Gondwana Research, vol. 75, p. 68-96.
Clifton R.L. (1944).- Ammonoids from Upper Cherry Canyon of Delaware Mountain Group in Texas.- AAPG Bulletin, Tulsa - OK, vol. 28, no. 11, p. 1644-1646.
Cooper G. (1953).- Sponges, Brachiopoda, Pelecypoda, and Scaphopoda. In: Cooper G., Dunbar C.O., Duncan H., Miller A.K. & Knight J.B. (eds.), Permian fauna at El Antimonio, western Sonora, Mexico.- Smithsonian Miscellaneous Collections, Washington - DC, vol. 119, no. 2, p. 21-80. URI: http://hdl.handle.net/10088/22872
Corona-Esquivel R. (1983).- Estratigraf�a de la regi�n de Olinal�-Tecocoyunca, noreste del Estado de Guerrero.- Revista Mexicana de Ciencias Geol�gicas, Quer�taro, vol. 5, no. 1, p. 17-24. URL: https://rmcg.geociencias.unam.mx/index.php/rmcg/article/view/1371
Corona-Esquivel R. (1985).- Geolog�a de la regi�n comprendida entre Olinal� y Huamuxtitl�n, Edo. de Guerrero.- Tesis de maestr�a, Universidad Nacional Aut�noma de M�xico, Ciudad de M�xico, 93 p. (7 Pls.). URI: https://hdl.handle.net/20.500.14330/TES01000773309
Dunbar C. (1944).- Part II. Permian and Pennsylvanian (?) fusulines. In: King R.E., Dunbar C.O., Cloud P.E. Jr & Miller A.K. (eds.), Geology and paleontology of the Permian area northwest of Las Delicias, southwestern Coahuila, Mexico.- Geological Society of America Special Paper, Boulder - CO, vol. 52, p. 35-48.
Ehiro M. (2008).- Two genera of Popanoceratidae (Permian Ammonoidea) from the South Kitakami Belt, Northeast Japan, with a note on the age of the Takakurayama Formation in the Abukuma Massif.- Bulletin of the Tohoku University Museum, Sendai, no. 8, p. 1-8. URL: https://www.museum.tohoku.ac.jp/pdf/press_info/bulletin/No08/bulletin_08_01.pdf
Ehiro M. (2022).- Middle Permian cephalopods of the Takakurayama Formation in the Yaguki area, the southwestern margin of the South Kitakami Belt, Northeast Japan.- Bulletin of the Tohoku University Museum, Sendai, no. 21, p. 21-38. URL: https://www.museum.tohoku.ac.jp/pdf/press_info/bulletin/No21/bulletin_21_02.pdf
Ehiro M. & Araki H. (1997).- 5. Permian cephalopods of Kurosawa, Kesennuma City in the Southern Kitakami Massif.- Paleontological Research, Tokyo, vol. 1, no. 1, p. 55-66. URL: https://www.palaeo-soc-japan.jp/publications/pr/Ehiro_Araki_1997.pdf
Ehiro M. & Misaki A. (2005).- Middle Permian ammonoids from the Kamiyasse-Imo district in the southern Kitakami Massif, northeast Japan.- Paleontological Research, Tokyo, vol. 9. no. 1, p. 1-14. URL: https://www.jstage.jst.go.jp/article/prpsj/9/1/9_1_1/_pdf
Flores de Dios A.B. & Buitr�n-S�nchez B.E. (1982).- Revisi�n y aportes a la estratigraf�a de la Monta�a de Guerrero.- Universidad Aut�noma de Guerrero, Serie T�cnico-Cient�fica, Chilpancingo, vol. 12, p. 1-28.
Furnish W.M. (1973).- Permian stage names. In: Logan A. & Hills L.V. (eds.), Permian and Triassic systems and their mutual boundary.- Canadian Society of Petroleum Geologists, Memoirs, vol. 2, p. 522-548.
Furnish W., Glenister B., Kullmann J. & Zhou Z. (2009).- Carboniferous and Permian Ammonoidea (Goniatitida and Prolecanitida). In: Selden P.A. (ed.), Treatise on Invertebrate Paleontology, Part L, Mollusca 4 (revised version), vol. 2.- Geological Society of America, Boulder - CO; University of Kansas Press, Lawrence - KS, Ch. 4, p. 158-192. DOI: 10.17161/dt.v0i0.5274
Garibay-Romero L.M. & Torres R. (1998).- Trilobite p�rmico de la Formaci�n Olinal� (Los Arcos), en el estado de Guerrero. In: VI Congreso Nacional de Paleontolog�a.- Sociedad Mexicana de Paleontolog�a, Memorias, Ciudad de M�xico, Res�menes, p. 23.
Garibay-Romero L.M. (2000).- Nuevas amonitas p�rmicos de la Formaci�n Olinal�-Los Arcos, Olinal�, Gro., M�xico (resumen). In: II Reuni�n Nacional de Ciencias de la Tierra, Puerto Vallarta.- GEOS, Ensenada, vol. 20, no. 3, p. 299. URL: https://www.ugm.org.mx/publicaciones/geos/pdf/geos00-3/PALEO00-3.pdf
Gemmellaro G. (1887).- La fauna dei calcari con Fusulina della valle del fiume Sosionella provincia di Parlermo.- Giornale di Scienze Naturali ed Economiche, Palermo, vol. 19, fasc. I, p. 1-106 (X Pls.).
Girty G.H. (1902).- The Upper Permian in western Texas.- American Journal of Science, vol. 4, no. 83, article XXXV, p. 363-368. DOI: 10.2475/ajs.s4-14.83.363
Girty G.H. (1908).- The Guadalupian Fauna.- U.S. Geological Survey, Professional Paper 58, Washington - DC, 651 p. DOI: 10.3133/pp58
Glenister B.F. & Furnish W.M. (1961).- The Permian ammonoids of Australia.- Journal of Paleontology, Cambridge (UK), vol. 35, no. 4, p. 673-736.
Glenister B.F. & Furnish W.M. (1988).- Patterns in stratigraphic distribution of Popanocerataceae, Permian ammonoids.- Senckenbergiana lethaea, vol. 69, nos. 1/2, p. 43-71.
Glenister B.F., Boyd D.W., Furnish W.M., Grant R.E., Harris M.T., Kozur H., Lambert L.L., Nassichuk W.W, Newel N.D., Pray L.C., Spinosa C., Wardlaw B.R., Wilde G.L. & Yancey T.E. (1992).- The Guadalupian: Proposed international standard for a Middle Permian Series.- International Geology Review, vol. 34, no. 9, p. 857-888.
Glenister B.F., Wardlaw B.R., Lambert L.L., Spinosa C., Bowring S.A. & Erwin D.H. (1999).- Proposal of Guadalupian and Component Roadian, Wordian and Capitanian Stages as International Standards for the Middle Permian Series.- Permophiles, Boise - ID, no. 34, p. 3-11.
Gonz�lez-Arreola C., Villase�or-Mart�nez A. & Corona-Esquivel R. (1994).- Permian fauna of the Los Arcos Formation, municipality of Olinal�, state of Guerrero, Mexico.- Revista Mexicana de Ciencias Geol�gicas, Quer�taro, vol. 11, no. 2, p. 214-221.
Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (2020).- Geologic Time Scale 2020.- Elsevier, Amsterdam, vol. 2, p. 565- 1357.
Grant R.E., Wardlaw B.R. & Rohr D.M. (2000).- Guadalupian studies in West Texas. In: Wardlaw B.R., Grant R.E. & Rohr D.M. (eds.), Guadalupian Symposium.- Smithsonian Contributions to Earth Sciences, Washington - DC, vol. 32, p. 1-4.
Guerrero-S�nchez M. & Quiroz-Barroso S.A. (2013).- Bivalvos guadalupianos (P�rmico medio) de la Formaci�n Olinal�, Guerrero, M�xico.- Bolet�n de la Sociedad Geol�gica Mexicana, Ciudad de M�xico, vol. 65, no. 3, p. 533-551. URL: 10.18268/BSGM2013v65n3a8
Guti�rrez-Quinto M.P. (2002, unpublished).- An�lisis de facies y microfacies del miembro carbonatado de la Formaci�n Olinal� (P�rmico), Estado de Guerrero: Taxco, Guerrero, M�xico.- Tesis profesional, Universidad Nacional Aut�noma de M�xico, Ciudad de M�xico, 109 p.
Henderson C.M. & Shen S.Z. (2020).- The Permian Period. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale 2020.- Elsevier, Amsterdam, vol. 2, p. 875-902.
Huang H., Shi Y.-K. & Jin X.-C. (2017).- Permian (Guadalupian) fusulinids of Bawei Section in Baoshan Block, western Yunnan, China: Biostratigraphy, facies distribution and paleogeographic discussion.- Palaeoworld, vol. 26, no. 1, p. 95-114.
International Commission on Stratigraphy (2026).- International Chronostratigraphic Chart.- URL: https://stratigraphy.org/chart/
Jin Y.G., Wardlaw B.R., Glenister B.F. & Kotlyar G.V. (1997).- Permian chronostratigraphic subdivisions.- Episodes, Seoul, vol. 20, no. 1, p. 10-15. DOI: 10.18814/epiiugs/1997/v20i1/003
Ju�rez-Arriaga E. & Murillo-Mu�eton G. (2020).- Arquitectura estratigr�fica, ambientes de dep�sito y geocronolog�a de la Formaci�n Olinal� (P�rmico tard�o), noreste de Guerrero, M�xico.- Revista Mexicana de Ciencias Geol�gicas, Quer�taro, vol. 37, no. 3, p. 179-196. DOI: 10.22201/cgeo.20072902e.2020.3.1576
King R.E. (1944).- Part I: Geology. In: King R.E., Dunbar C.O., Cloud P.E. Jr & Miller A.K. (eds.), Geology and paleontology of the Permian area northwest of Las Delicias, southwestern Coahuila, Mexico.- Geological Society of America Special Paper, Boulder - CO, vol. 52, p. 3-35.
Klug C., Korn D., Landman N.H., Tanabe K., De Baets K. & Naglik C. (2015).- Describing ammonoid conchs. In: Klug C., Korn D., De Baets K., Kruta I. & Mapes R.H. (eds.), Ammonoid paleobiology: From macroevolution to paleogeography.- Topics in Geobiology, vol. 44, p. 3-24.
Korn D. (2010).- A key for the description of Palaeozoic ammonoids.- Fossil Record, G�ttingen, vol. 13, no. 1, p. 5-12. DOI: 10.1002/mmng.200900008
Kullmann J., Korn D. & Petersen M.S. (2000).- GONIAT database system, version 2.90.- Geological Institute of T�bingen. URL: https://www.goniat.org/
Lambert L.L., Lehrmann D.J., Harris M.T., Wardlaw B.R., Grant R.E. & Rohr D.M. (2000).- Correlation of the Road Canyon and Cutoff formations, West Texas, and its relevance to establishing an international Middle Permian (Guadalupian) series in The Guadalupian Symposium.- Smithsonian Contributions to Earth Sciences, Washington - DC, vol. 32, p. 153-173.
Lambert L.L., Wardlaw B.R., Nestell M.K. & Nestell G.P. (2002).- Latest Guadalupian (Middle Permian) conodonts and foraminifers from West Texas.- Micropaleontology, New York - NY, vol. 48, no. 4, p. 343-364.
Lara-Pe�a R.A., Navas-Parejo P. & Torres-Mart�nez M.A. (2021).- Permian autochthony of northwestern Mexico based on conodont paleogeographic relationships with southwestern Laurentia.- Newsletters on Stratigraphy, Stuttgart, vol. 54, no. 3, p. 363-376.
Leonova T.B. (1999).- Stages in evolution and biogeography of Permian ammonoids.- Stratigraphy and Geological Correlation, vol. 7, no. 6, p. 568-580.
Leonova T.B. (2002).- Permian ammonoids, classification and phylogeny.- Paleontologicheskii Zhurnal, Moscow, vol. 36, supplement 1, p. S1-S114. URI: https://repository.geologyscience.ru/handle/123456789/29103
Leonova T.B. (2009).- Ammonoid evolution in marine ecosystems prior to the Permian-Triassic crisis.- Paleontological Journal, Moscow, vol. 43, p. 858-865.
Leonova T.B. (2010).- Revision of the Permian Ammonoid Family Cyclolobidae.- Paleontological Journal, Moscow, vol. 44, no. 3, p. 267-274.
Leonova T.B. (2011).- Permian ammonoids: Biostratigraphic, biogeographical, and ecological analysis.- Paleontological Journal, Moscow, vol. 45, no. 10, p. 1206-1312.
Leonova T.B. (2016).- Major trends in the evolution of Permian ammonoids.- Paleontological Journal, Moscow, vol. 50, p. 131-140.
Leonova T.B. (2017).- Evolution of Palaeozoic ammonoid sutures.- Invertebrate Zoology, Moscow, vol. 14. no. 1, p. 27-31. DOI: 10.15298/invertzool.14.1.05
Leonova T.B. (2018).- Permian ammonoid biostratigraphy. In: Lucas S.G. & Shen S.Z. (eds.), The Permian timescale.- Geological Society, London, Special Publications, vol. 450, no. 1, p. 185-203.
Leonova T.B. & Boiko M.S. (2015).- Phylogeny of the Late Paleozoic superfamily Adrianitoidea Schindewolf (Ammonoidea).- Paleontological Journal, Moscow, vol. 49, no. 6, p. 584-594.
Miller A.K. (1938).- Comparison of Permian ammonoid zones of Soviet Russia with those of North America.- AAPG Bulletin, Tulsa - OK, vol. 22, no. 8, p. 1014-1019.
Miller A. (1944).- Part IV: Permian cephalopods. In: King R.E., Dunbar C.O., Cloud P.E. Jr & Miller A.K. (eds.), Geology and paleontology of the Permian area Northwest of Las Delicias, Southwestern Coahuila, Mexico.- Geological Society of America Special Paper, Boulder - CO, vol. 52, p. 71-128.
Miller A. & Furnish W. (1940).- Permian ammonoids of the Guadalupe Mountain region and adjacent areas.- Geological Society of America Special Paper, Boulder - CO, vol. 26, 242 p.
Mor�n-Cuevas E. (2007, unpublished).- Descripci�n de facies e interpretaci�n deposicional del miembro cl�stico de la Formacion Olinal� (P�rmico), Estado de Guerrero.- Tesis profesional, Escuela Regional de Ciencias de Tierra, Universidad Aut�noma de Guerrero, Chilpancingo, 102 p.
Murphy M.A. & Salvador A. (eds., 2000).- International Subcommission on Stratigraphic Classification of IUGS International Commission on Stratigraphy: International stratigraphic guide - An abridged version.-GeoArabia, Manama, vol. 5, no. 2, p. 231-266. DOI: 10.2113/geoarabia0502231
Nassichuk W. (1977).- Permian ammonoids from the Cache Creek Group in Western Canada.- Journal of Paleontology, Cambridge (UK), vol. 51, no. 3, p. 557-590.
Nestell M.K., Nestell G.P. & Wardlaw B.R. (2019).- Integrated fusulinid, conodont, and radiolarian biostratigraphy of the Guadalupian (Middle Permian) in the Permian Basin region, USA. In: Ruppel S.C. (ed.), Anatomy of a Paleozoic basin: The Permian Basin, USA (vol. 1, ch. 9).- Bureau of Economic Geology Report of Investigations, The University of Texas at Austin, no. 285; AAPG Memoir, Tulsa - OK, no. 118, p. 251-291.
Plummer F. & Scott G. (1937).- Upper Paleozoic ammonites in Texas. In: Sellards E.H. (ed.), The geology of Texas, vol. III. Upper Paleozoic Ammonites and Fusulinids, part 1.- The University Texas Bulletin, Austin - TX, no. 3701, p. 11-516.
Ortega-Guti�rrez F., El�as-Herrera M., Reyes-Salas M., Mac�as-Romero C. & L�pez R. (1999).- Late Ordovician-Early Silurian continental collisional orogeny in Southern Mexico and its bearing on Gondwana-Laurentia connections.- Geology, Boulder - CO, vol. 27, no. 8, p. 719-722.
Ruzhencev V.E. (1950).- Verkhnekamennougol'nye ammonity Urala (Upper Carboniferous ammonites of the Urals).- Trudy Komi filiala, Akademii Nauk SSSR, Syktyvkar, vol. 29, 220 p.
Ruzhencev V.E. (1955).- Main stratigraphic assemblages of ammonoids of the Permian system.- Izvestiya Akademii Nauk SSSR (Seriya Biologicheskaya), Moscow, no. 4, p. 120-132.
Ruzhencev V.E. (1976).- Late Permian ammonoids in the Far East.- Paleontologicheskii Zhurnal, Moscow, vol. 3, p. 36-50.
Shen S., Zhang H., Zhang Y., Yuan D., Chen B., He W., Mu L., Lin W., Wang W., Chen J., Wu Q., Cao C., Wang Y. & Wang X. (2019).- Permian integrative stratigraphy and timescale of China.- Science China Earth Sciences, Beijing, vol. 62, p. 154-188.
Shen S.Z., Yuan D.X., Henderson C.M., Wu Q., Zhang Y.C., Zhang H., Mu L., Ramezani J., Wang X., Lambert L., Erwin D., Hearst J., Xiang L., Chen B., Fan J., Wang Y., Wang W., Qi Y., Chen J., Qie W. & Wang T.T. (2020).- Progress, problems and prospects: An overview of the Guadalupian Series of South China and North America.- Earth-Science Reviews, vol. 211, article 103412, 26 p.
Shumard B.F. (1858).- Notice of new fossils from the Permian strata of New Mexico and Texas, collected by Dr. George G. Shumard, geologist of the United States Government expedition for obtaining water by means of artesian wells along the 32nd parallel, under the direction of Capt John Pope, U.S. Corps Top. Eng.- Transactions of the Academy of Science of St. Louis, St. Louis - MO, vol. I, p. 290-297. URL: https://www.biodiversitylibrary.org/page/30339587
Smith A.N. (2012).- New ammonoid localities from the southern Delaware Mountains and Seven Heart Gap, Culberson County, west Texas.- Thesis Master of Science in Geology, The University of Texas at San Antonio, 54 p. URL: https://hdl.handle.net/20.500.12588/5755
Spinosa C., Furnish W. & Glenister B. (1975).- The Xenodiscidae, Permian ceratitoid ammonoids.- Journal of Paleontology, Cambridge (UK), vol. 49, no. 2, p. 239-283.
Talavera-Mendoza O., Ruiz J., Gehrels G.E., Meza-Figueroa D.M., Vega-Granillo R. & Campa-Uranga M.F. (2005).- U-Pb geochronology of the Acatl�n Complex and implications for the Paleozoic paleogeography and tectonic evolution of southern Mexico.- Earth and Planetary Science Letters, vol. 235, no. 3-4, p. 682-Tazawa.
Tazawa J.I., Fujikawa M., Zakharov Y.D. & Hasegawa S. (2005).- Middle Permian ammonoids from the Takakurayama area, Abukuma Mountains, northeast Japan, and their stratigraphical significance.- Science reports of Niigata University, no. 20, p. 15-27. URL: https://museum.sc.niigata-u.ac.jp/reports/2005/report2005_02.pdf
Vachard D., Oviedo A., Flores de Dios A., Malpica R., Brunner P., Guerrero-Suastegui M. & Buitr�n-S�nchez B. (1993).- Barranca d'Olinal� (Guerrero) : Une coupe de r�ference pour le Permien du Mexique central ; �tude pr�liminaire.- Annales de la Societ� G�ologique du Nord, Lille, vol. 2, p. 155-162.
Vachard D., Flores de Dios A. & Buitr�n-S�nchez B.E. (1997).- Sur une nouvelle localit� � Fusulines du Wordien (Permien sup�rieur) du Mexique ; cons�quences pal�og�ographiques.- Geobios, Villeurbanne, vol. 30, no. 3, p. 361-370.
Vachard D., Flores de Dios A. & Buitr�n-S�nchez B.E. (2004).- Guadalupian and Lopingian (Middle and Late Permian) deposits from Mexico and Guatemala, a review with new data.- Geobios, Villeurbanne, vol. 37, no. 1, p. 99-115.
Vega-Hern�ndez E.D. (2011).- An�lisis petrogr�fico y diagen�tico del Miembro carbonatado de la Formaci�n Olinal�, del P�rmico medio, en la regi�n de la Monta�a de Guerrero.- Tesis profesional, Escuela Regional de Ciencias de Tierra, Universidad Aut�noma de Guerrero, Chilpancingo, 68 p.
Villase�or-Mart�nez A.B., Mart�nez-Cort�s A. & Contreras B. (1987).- Bioestratigraf�a del Paleozoico Superior de San Salvador Patlanoaya, Puebla, M�xico.- Revista de la Sociedad Mexicana de Paleontolog�a, Quer�taro, vol. 1, no. 1, p. 396-417.
Wahlman G.P. & Nestell M.K. (2024).- Middle Permian (Guadalupian) Fusulinids from subsurface Midland Basin, West Texas, USA, including paleoenvironmental microspheric Parafusulina.- Journal of Foraminiferal Research, Glen Allen - VA, vol. 54, no. 2, p. 117-142.
Wilde G.L. (1955).- Permian fusulinids of the Guadalupe Mountains. In: Permian Field Conference, October 1955.- Permian Basin Section, SEPM, Midland - TX, Publication 55-1, p. 59-62.
Wu Q., Ramezani J., Zhang H., Wang T.T., Yuan D.X., Mu L., Zhang Y., Li X. & Shen S.Z. (2017).- Calibrating the Guadalupian Series (Middle Permian) of South China.- Palæogeography, Palæoclimatology, Palæoecology, vol. 466, p. 361-372.
Yang Z.D. & Yancey T.E. (2000).- Fusulinid biostratigraphy and paleontology of the Middle Permian (Guadalupian) strata of the Glass Mountains and Del Norte Mountains, West Texas. In: Wardlaw B.R., Grant R.E. & Rohr D.M. (eds.), Guadalupian Symposium.- Smithsonian Contributions to Earth Sciences, Washington - DC, vol. 32, p. 185-259.
Zhang Y.C. & Wang Y. (2018).- Permian fusuline biostratigraphy. In: Lucas S.G. & Shen S.Z. (eds.), The Permian timescale.- Geological Society, London, Special Publications, vol. 450, p. 253-288.
Zhang S.H., Yuan D.X. & Shen S.Z. (2024).- Global correlation of the Guadalupian-Lopingian transition and associated events using a quantitative conodont-based biostratigraphic scheme.- Palæogeography, Palæoclimatology, Palæoecology, vol. 655, article 112502, 13 p.
Zhou Z. (2017).- Permian basinal ammonoid sequence in Nanpanjiang area of South China possible overlap between basinal Guadalupian and platform-based Lopingian.- Journal of Paleontology, Cambridge (UK), vol. 91, p. 1-95.
For the fieldwork bed number column, the bold letters indicate the bed number of the section where the ammonoid was collected, while the regular text indicates the specimen number within that bed. For example, Nemo2-4 corresponds to specimen 4 from level Nemo 2 of the Nemotitl�n section.
| Collection number | Fieldwork bed number | Identification | Figured |
| Oli1 | |||
| 14378 | Oli1-1 | Roadoceras rodense | |
| 14379 | Oli1-2 | Roadoceras rodense | |
| 14380 | Oli1-3 | Roadoceras rodense | |
| 14381 | Oli1-4 | Roadoceras rodense | |
| 14382 | Oli1-5 | Roadoceras rodense | |
| 14383 | Oli1-6 | Roadoceras rodense | |
| 14384 | Oli1-7 | Roadoceras rodense | |
| 14385 | Oli1-8 | Roadoceras rodense | |
| 14386 | Oli1-9 | Roadoceras rodense | |
| 14387 | Oli1-10 | Roadoceras rodense | |
| 14388 | Oli1-11 | Roadoceras rodense | |
| 14389 | Oli1-12 | Roadoceras rodense | |
| 14390 | Oli1-13 | Roadoceras rodense | |
| 14391 | Oli1-14 | Roadoceras rodense | |
| 14392 | Oli1-15 | Roadoceras rodense | |
| 14393 | Oli1-16 | Roadoceras rodense | |
| 14394 | Oli1-17 | Roadoceras rodense | |
| 14395 | Oli1-18 | Roadoceras rodense | |
| 14396 | Oli1-19 | Roadoceras rodense | |
| 14397 | Oli1-20 | Roadoceras rodense | |
| 14398 | Oli1-21 | Roadoceras rodense | |
| 14399 | Oli1-22 | Roadoceras rodense | |
| 14400 | Oli1-23 | Roadoceras rodense | |
| 14401 | Oli1-24 | Adrianites sp. |
Fig. 3.G |
| Oli2 | |||
| 14402 | Oli2-1 | Pseudagathiceras spinosum |
Fig. 3.C-D |
| 14403 | Oli2-2 | Roadoceras rodense | |
| 14404 | Oli2-3 | Roadoceras rodense |
Fig. 3.I |
| 14405 | Oli2-4 | Roadoceras rodense |
Fig. 3.J |
| 14406 | Oli2-5 | Roadoceras rodense | |
| 14407 | Oli2-6 | Indeterminate | |
| Te2 | |||
| 14408 | Te2-1 | Paraceltites elegans |
Fig. 2.K
|
| 14409 | Te2-2 | Paraceltites elegans |
Fig. 2.J
|
| 14410 | Te2-3 | Paraceltites elegans | |
| 14411 | Te2-4 | Paraceltites elegans | |
| 14412 | Te2-5 | Paraceltites elegans | |
| 14413 | Te2-6 | Paraceltites elegans | |
| 14414 | Te2-7 | Paraceltites elegans | |
| 14415 | Te2-8 | Paraceltites elegans | |
| 14416 | Te2-9 | Paraceltites elegans | |
| 14417 | Te2-10 | Paraceltites elegans | |
| 14418 | Te2-11 | Paraceltites elegans | |
| 14419 | Te2-12 | Paraceltites elegans | |
| 14420 | Te2-13 | Paraceltites elegans | |
| 14421 | Te2-14 | Paraceltites elegans | |
| 14422 | Te2-15 | Paraceltites elegans | |
| 14423 | Te2-16 | Paraceltites elegans | |
| 14424 | Te2-17 | Paraceltites elegans | |
| 14425 | Te2-18 | Pseudagathiceras spinosum | |
| 14426 | Te2-19 | Agathiceras sp. |
Fig. 3.E |
| 14427 | Te2-20 | Agathiceras sp. |
Fig. 3.F |
| 14428 | Te2-21 | Agathiceras sp. | |
| 14429 | Te2-22 | Metacrimites sp. | |
| 14430 | Te2-23 | Agathiceras sp. | |
| 14431 | Te2-24 | Agathiceras sp. | |
| 14432 | Te2-25 | Agathiceras sp. | |
| 14433 | Te2-26 | Agathiceras sp. | |
| 14434 | Te2-27 | Indeterminate | |
| 14435 | Te2-28 | Metacrimites sp. | |
| 14436 | Te2-29 | Epithalassoceras sp. | |
| 14437 | Te2-30 | Roadoceras roadense | |
| 14438 | Te2-31 | Paraceltites elegans | |
| 14439 | Te2-32 | Propinacoceras sp. |
Fig. 3.A |
| 14440 | Te2-33 | Epithalassoceras sp. |
Fig. 3.H |
| 14441 | Te2-34 | Metacrimites sp. |
Fig. 3.K |
| Nemo2 | |||
| 14442 | Nemo2-1 | Indeterminate | |
| 14443 | Nemo2-2 | Paraceltites elegans |
Fig. 2.I
|
| 14444 | Nemo2-3 | Roadoceras roadense | |
| 14445 | Nemo2-4 | Roadoceras roadense | |
| 14446 | Nemo2-5 | Roadoceras roadense | |
| 14447 | Nemo2-6 | Roadoceras roadense | |
| 14448 | Nemo2-7 | Roadoceras roadense | |
| Nemo3 | |||
| 14449 | Nemo3-1 | Waagenoceras cf. W. dieneri |
Fig. 2.G
|
| 14450 | Nemo3-2 | Waagenoceras cf. W. dieneri |
Fig. 2.E
|
| 14451 | Nemo3-3 | Mexicoceras sp. |
Fig. 2.C-D
|
| 14452 | Nemo3-4 | Waagenoceras cf. W. dieneri | |
| 14453 | Nemo3-5 | Waagenoceras cf. W. dieneri | |
| 14454 | Nemo3-6 | Nautiloid | |
| 14455 | Nemo3-7 | Nautiloid | |
| 14456 | Nemo3-8 | Waagenoceras cf. W. dieneri | |
| 14457 | Nemo3-9 | Waagenoceras cf. W. dieneri | |
| 14458 | Nemo3-10 | Waagenoceras cf. W. dieneri | |
| 14459 | Nemo3-11 | Coahuiloceras sp. |
Fig. 6 |
| 14460 | Nemo3-12 | Stacheoceras sp. | |
| 14461 | Nemo3-13 | Stacheoceras sp. | |
| 14462 | Nemo3-14 | Stacheoceras sp. | |
| 14463 | Nemo3-15 | Stacheoceras sp. | |
| 14464 | Nemo3-16 | Stacheoceras sp. | |
| 14465 | Nemo3-17 | Stacheoceras sp. | |
| 14466 | Nemo3-18 | Stacheoceras sp. | |
| 14467 | Nemo3-19 | Stacheoceras sp. | |
| 14468 | Nemo3-20 | Propinacoceras beyrichi |
Fig. 3.B |
| 14469 | Nemo3-21 | Coahuiloceras sp. | |
| 14470 | Nemo3-22 | Coahuiloceras sp. | |
| 14471 | Nemo3-23 | Waagenoceras cf. W. girtyi |
Fig. 2.H
|
| 14472 | Nemo3-24 | Waagenoceras cf. W. girtyi | |
| 14473 | Nemo3-25 | Indeterminate | |
| 14474 | Nemo3-26 | Indeterminate | |
| Coyo3 | |||
| 14475 | Coyo3-1 | Waagenoceras cf. W. dieneri | |
| 14476 | Coyo3-2 | Waagenoceras cf. W. dieneri | |
| 14477 | Coyo3-3 | Waagenoceras cf. W. dieneri | |
| 14478 | Coyo3-4 | Waagenoceras cf. W. dieneri | |
| 14479 | Coyo3-5 | Waagenoceras cf. W. dieneri | |
| 14480 | Coyo3-6 | Waagenoceras cf. W. dieneri | |
| 14481 | Coyo3-7 | Waagenoceras cf. W. dieneri |
Fig. 2.F
|
| 14482 | Coyo3-8 | Stacheoceras sp. | |
| 14483 | Coyo3-9 | Stacheoceras sp. | |
| 14484 | Coyo3-10 | Stacheoceras sp. | |
| 14485 | Coyo3-11 | Stacheoceras sp. | |
| 14486 | Coyo3-12 | Stacheoceras sp. | |
| 14487 | Coyo3-13 | Stacheoceras sp. | |
| 14488 | Coyo3-14 | Stacheoceras sp. | |
| 14489 | Coyo3-15 | Stacheoceras sp. |
Fig. 2.B
|
| 14490 | Coyo3-16 | Stacheoceras sp. | |
| 14491 | Coyo3-17 | Stacheoceras sp. | |
| 14492 | Coyo3-18 | Stacheoceras sp. | |
| 14493 | Coyo3-19 | Stacheoceras sp. | |
| 14494 | Coyo3-20 | Roadoceras roadense | |
| 14495 | Coyo3-21 | Roadoceras roadense | |
| 14496 | Coyo3-22 | Stacheoceras sp. | |
| 14497 | Coyo3-23 | Stacheoceras sp. |
Fig. 2.A
|