◄ Carnets Geol. 26 (3) ►
![]()
Outline:
[1. Introduction]
[2. Regional geographical and geological settings]
[3. Stratigraphy of the Saint-Montan area and correlations]
[4. Systematic palaeontology]
[5. Discussion]
[6. Evolutionary aspects]
[7. Conclusions]
and ... [Bibliographic references]
Aix Marseille Université, CEREGE, Place Victor Hugo, Marseille, Cedex 03, 13331 (France)
Sorbonne Université, MNHN, CNRS, Centre de Recherche en Paléontologie- Paris, CR2P, F-75252 Paris (France)
Aix Marseille Université, CEREGE, Place Victor Hugo, Marseille, Cedex 03, 13331 (France)
Groupement d'Intéręt Paléontologique, Science et Exposition, 60 Bd Georges Richard, 83000, Toulon (France)
Published online in final form (pdf) on January 21, 2026
DOI:
10.2110/carnets.2026.2603
![]()
[Editor: Bruno
R.C. Granier; language editor: Robert W. Scott]
![]()
Upper Barremian-lower Aptian Urgonian-type platform carbonates from Ardčche yield a rich rudist fauna belonging to the families Requieniidae, Monopleuridae, Caprinidae, and Polyconitidae. The upper Barremian assemblage from the Ardčche river gorges consists of eighteen species, it conforms to the Rustrel fauna of the Monts-de-Vaucluse but contains a new genus Valclusella, and a new species of Bicornucopina, a genus unknown so far in Western Europe, whereas Mathesia darderi is identified for the first time in SE France. We define the "Saint-Montan limestones", of early Aptian age, as the uppermost member of the Urgonian Formation in this region. The corresponding Saint-Montan fauna is described for the first time: it contains twenty-five rudist species, of which eight are new and seventeen are inherited from the antecedent late Barremian Orgon and Rustrel fossil type localities. Of the eight new species, two are ascribed to earlier genera, i.e., Homopleura parva and Bicornucopina acuminata, three are assigned to a new genus, Myostyla ardescensis, while Valclusella valclusensis and Valclusella biconvexa preexisted in the late Barremian. Aside from inherited taxa, e.g., Pachytraga paradoxa, Praecaprina varians-gaudryi, Caprina douvillei, important novelties regard the Caprinidae, characterized by the appearance of probable new species of Pachytraga, P. aff. paradoxa, Offneria, O. aff. rhodanica, and three new species of Praecaprina: Pr. tubifera n. sp., Pr. paquieri n. sp., and Pr. vacua n. sp. The early Aptian p.p., therefore, coincides with the acme of the genus Praecaprina, SE France being unique in this respect. In the Saint-Montan fauna, up to about nine species from the Rustrel fauna are missing. The Saint-Montan rudist assemblage is unique, thus far, in SE France, even as the study of its composition is still in progress. It has a West European character, similar to that of its antecedent late Barremian one, and the succeeding late early Aptian Villaroya de los Pinares fauna from Spain, but the three have distinctive taxonomic compositions. The Saint-Montan stratigraphic succession is framed by two drowning boundary events. The taxonomic composition of its rudist assemblage fills a gap between the late Barremian and the late early Aptian faunas, and represents a distinctive episode in the Barremian-lower Aptian Urgonian-type rudist sequence of southern France and Spain, and possesses a significant biostratigraphic potential.
• Rudist bivalves;
• upper Barremian-lower Aptian;
• taxonomy;
• Ardčche;
• south-east France
Masse J.-P., Rineau V., Fenerci-Masse M. & Frau C. (2026).- Upper Barremian-lower Aptian rudist faunas of Urgonian-type platform formations from Ardčche (southeastern France).- Carnets Geol., Madrid, vol. 26, no. 3, p. 51-87. DOI: 10.2110/carnets.2026.2603
Les rudistes des formations urgoniennes du Barrémien supérieur et de l'Aptien inférieur d'Ardčche (SE de la France).- Les formations carbonatées urgoniennes du Barrémien supérieur-Aptien inférieur d'Ardčche livrent une riche faune de rudistes appartenant aux Requieniidae, Monopleuridae, Caprinidae et Polyconitidae. L'assemblage du Barrémien supérieur, des gorges de l'Ardčche, est semblable ŕ celui de la faune de Rustrel des Monts-de-Vaucluse. Il comporte dix huit espčces, un genre nouveau Valclusella, une espčce nouvelle de Bicornucopina, genre inconnu jusqu'ici sur la marge ouest-européenne, tandis que Mathesia darderi est identifié pour la premičre fois dans le SE de la France. Les "Calcaires de Saint-Montan" d'âge aptien inférieur constituent le membre le plus élevé de la formation urgonienne régionale. La "faune de Saint-Montan", inédite, comprend vingt cinq espčces dont huit nouvelles, et dix sept héritées des localités fossilifčres barrémiennes, antécédentes d'Orgon et de Rustrel. Parmi les espčces nouvelles deux appartiennent ŕ des genres connus, ce sont Homopleura parva et Bicornucopina acuminata, trois relčvent de genres nouveaux : Myostyla ardescensis, forme exclusive, Valclusella valclusensis et Valclusella biconvexa déjŕ présentes au Barrémien supérieur. Ŕ côté des formes héritées, e.g., Pachytraga paradoxa, Praecaprina varians-gaudryi, Caprina douvillei, d'importantes innovations concernent les Caprinidae, caracterisés par l'apparition de probables espčces originales de Pachytraga, P. aff. paradoxa, Offneria, O. aff. rhodanica, et trois espčces de Praecaprina : Pr. tubifera n. sp., Pr. paquieri n. sp. et Pr. vacua n. sp. L'Aptien inferieur est donc marqué par l'acme du genre Praecaprina, le SE de la France jouant de ce point de vue un rôle privilégié. Dans le materiel étudié neuf espčces de la faune de Rustrel manquent. L'assemblage taxonomique des couches de Saint-Montan, dont le present article donne un aperçu incomplet, est unique et possčde comme la faune antécédente de Rustrel et celle plus récente de Villaroya de los Pinarčs d'Espagne, un caractčre ouest européen, mais ces trois assemblages ont des compositions differentes. La série de Saint-Montan dont l'inventaire taxonomique comble une lacune entre les deux faunes précédentes, est encadrée par deux discontinuités sédimentaires de drowning. L'assemblage fossilifčre de cette localité identifie un épisode distinct et singulier de la séquence ŕ rudistes des formations urgoniennes d'âge Barrémien-Aptien du sud de la France et d'Espagne. Il possčde un potentiel biostratigraphique important.
• rudistes ;
• bivalves ;
• Barrémien supérieur-Aptien inférieur ;
• taxinomie ;
• Ardčche ;
• sud-est de la France
Following the recent threefold stratigraphic distribution established in SE France for late Barremian rudist assemblages associated with Urgonian successions, the existence of early Aptian forms in this region has been denied, and evidence for contemporaneous faunas is mostly derived from Spain and the French Pyrenées (Masse, 1996; Skelton et al., 2010; Gili et al., 2016; Bilotte et al., 2017; Masse et al., 2020).
Historically, the recognition of a lower Aptian rudist fauna in the Urgonian platform successions of SE France was proposed by Paquier (1903, 1905), who's conclusion was grounded on two key aspects:
His stratigraphic interpretation of the Urgonian succession of the Subalpine Chains follows Lory's (1846) tripartite division of the formation. This classification comprises a lower Urgonian division assigned to the upper Barremian and an upper Urgonian division of early Aptian age. These formations are separated by a thin middle division, i.e., the "lower Orbitolina marls", aimed at correlating with the upper Barremian "Heteroceras marls" of the Vocontian basinal settings (Paquier, 1900).
The identification of a distinctive rudist assemblage, predominantly composed of Caprinidae, was attributed to the upper Urgonian Formation, and it was consequently assigned to the early Aptian (Paquier, 1896, 1905).
These views were followed and applied to neighboring regions of SE France, namely in southern and northern Provence (Blanc & Masse, 1965; Masse, 1976) and Bas-Vivarais (Lafarge, 1978; Cotillon et al., 1979). The existence of a lower Aptian rudist assemblage, dominated by Caprinidae, in SE France was then accepted, complemented and developed by Masse (1993, 2003), Skelton and Masse (1998), and Masse and Fenerci-Masse (2011, 2013a, 2013b, 2013c), and was, for instance, followed by Chartrousse (1998), Léonide et al. (2012), Skelton et al. (2019), and Martinez-Rodriguez et al. (2025). But the following demonstrates that this model is no longer valid.
This study is supported by the reevaluation of Urgonian formations of northern Provence, as undertaken by Frau et al. (2018) and Tendil et al. (2018). This research established updated age constraints by incorporating pivotal ammonite findings beneath the U2 Member of Leenhardt (1883), which is characterized by the high abundance and diversity of caprinid rudists, and within the overlying U3 Member, mostly bioclastic with a cherty basal unit. The revised stratigraphic model of the north Provence Urgonian succession referred to the ammonite scale (Reboulet et al., 2014), suggests relocating the caprinid rudist fauna from the U2 Member to the latest Barremian (lower part of the Martelites sarasini ammonite zone). Meanwhile, the Barremian/Aptian boundary is placed in the lowermost part of the U3 member (see below in the section "correlations"). This member has yielded Deshayesites sp., Procheloniceras gr. dechauxi and Pseudohaploceras matheroni indicative of the Deshayesites oglanlensis and D. forbesi zones (Frau et al., 2018). The foregoing shows that the lateral stratigraphic equivalence between the caprinid-bearing unit U2 and the bioclastic cherty unit U3, ascribed to "channels", postulated by Clavel et al. (2013), is hardly acceptable.
Recent
investigations in the Ardčche region (Bas Vivarais domain) resulted in the
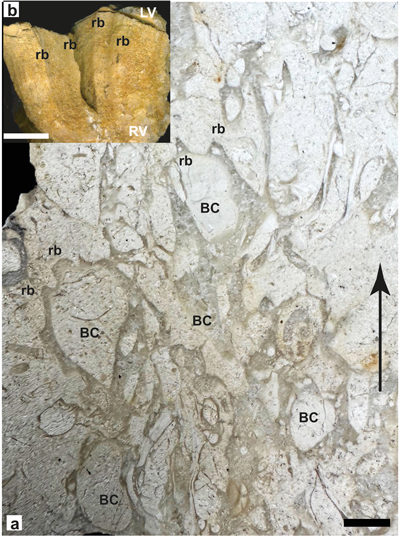
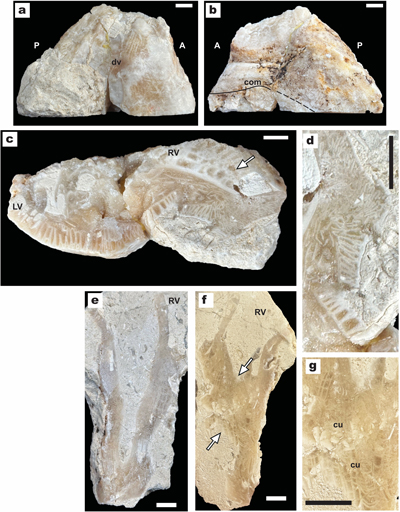
recognition of an atypical U3 member at Saint-Montan (Fig. 1 ![]() ), containing
rudists, first mentioned by Tendil et
al. (2018: Fig. 15). The primary objective of the present paper is to
investigate and characterize the rudist fauna associated with the lower Aptian
Saint-Montan limestones in its type locality and adjacent localities and to
decipher the analogies and differences with the antecedent late Barremian faunas
defined earlier: the Brouzet, Orgon, and Rustrel faunas (Masse et
al., 2020). Our study includes four rudist families: Requieniidae,
Monopleuridae, Polyconitidae, and Caprinidae. We also complement the inventory
of the Barremian Rustrel fauna after the study of geological sites from Ardčche.
), containing
rudists, first mentioned by Tendil et
al. (2018: Fig. 15). The primary objective of the present paper is to
investigate and characterize the rudist fauna associated with the lower Aptian
Saint-Montan limestones in its type locality and adjacent localities and to
decipher the analogies and differences with the antecedent late Barremian faunas
defined earlier: the Brouzet, Orgon, and Rustrel faunas (Masse et
al., 2020). Our study includes four rudist families: Requieniidae,
Monopleuridae, Polyconitidae, and Caprinidae. We also complement the inventory
of the Barremian Rustrel fauna after the study of geological sites from Ardčche.
|
|
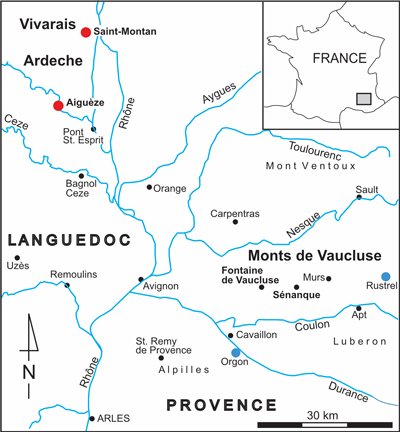
Figure 1:
Geographical
location of Barremian-lower Aptian Urgonian sites of Provence and Languedoc. The
key localities for rudist faunas are in red. Reference localities used for
comparisons are in blue. |
Previous studies on the Urgonian series of the Ardčche region (Bas-Vivarais) dealt with the stratigraphic insights of Lafarge (1978), Cotillon et al. (1979), Quesne (1996), which were subsequently updated by Clavel et al. (2014), Bastide (2014), Tendil et al. (2018), and Granier et al. (2021). Paleontological investigations regarding specific groups were undertaken by Paquier (1903, 1905), Löser and Ferry (2006) with timely contributions by Fenerci-Masse and Masse (2010), or are associated with the above mentioned publications.
In Ardčche
the type locality for early Aptian rudists is Saint-Montan and also three other
fossiliferous localities were investigated: Aigučze, Trescouvieux and Cité du
barrage. From this region, two sampling sites from the Ardčche river gorges,
with late Barremian rudists: Combe des Pins and Combe de Gournier (Frau et
al., unpublished) (Fig. 2 ![]() ) were also prospected. In the Monts de Vaucluse,
we addressed a single fossil-bearing locality, les Chaux de la Tour quarry,
which is a portion of the Urgonian succession of Fontaine de Vaucluse,
corresponding with the U2 member (Masse, 1976; Léonide et
al., 2012).
) were also prospected. In the Monts de Vaucluse,
we addressed a single fossil-bearing locality, les Chaux de la Tour quarry,
which is a portion of the Urgonian succession of Fontaine de Vaucluse,
corresponding with the U2 member (Masse, 1976; Léonide et
al., 2012).
|
|
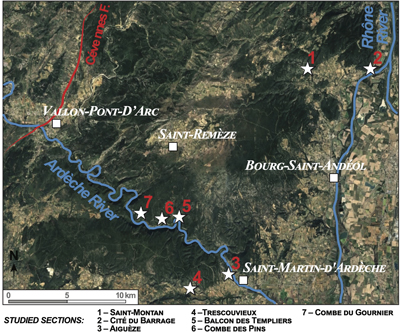
Figure 2:
Spatial distribution of the investigated lower Aptian (sites 1-4) and upper Barremian (sites
5- 6) fossil bearing sections in the Ardčche
area. |
The
initial stratigraphic description of the Saint-Montan section (Fig. 2 ![]() , site 1)
was presented by Lafarge (1978), discussed by Cotillon et
al. (1979) and Masse and Fenerci-Masse
(2011), then
subsequently updated by Clavel et al. (2014) and Tendil et
al. (2018). The Urgonian succession consists of the following five SM units sensu
Lafarge (1978), each bounded by stratigraphic discontinuities (Fig. 3
, site 1)
was presented by Lafarge (1978), discussed by Cotillon et
al. (1979) and Masse and Fenerci-Masse
(2011), then
subsequently updated by Clavel et al. (2014) and Tendil et
al. (2018). The Urgonian succession consists of the following five SM units sensu
Lafarge (1978), each bounded by stratigraphic discontinuities (Fig. 3 ![]() ),
and its dating refers to the Reboulet et
al. (2014) ammonite scale.
),
and its dating refers to the Reboulet et
al. (2014) ammonite scale.
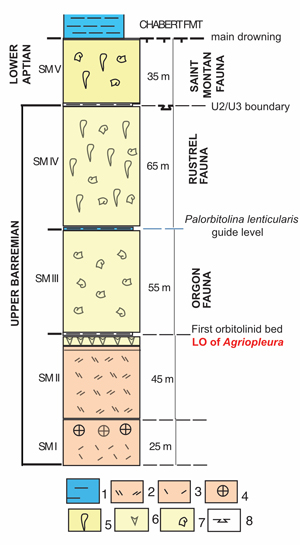
Unit SM I (25 m thick), consists of coral-bearing beds, including chaetetids, scleractinians with some oysters, capped by a hard-ground. Clavel et al. (2014) attributed these beds to the T. vandenheckei zone.
Unit SM II (45 m thick) consists of biocalcarenites with orbitolinids, dasycladale algae and echinoid fragments. The upper part is marked by coral beds, overlain by rudist beds. The uppermost rudist beds with Agriopleura marticensis are capped by a discontinuity and overlain by a rudist-bearing marly bed. This marly bed, yielding the orbitolinids Palaeodictyoconus actinostoma and Paracoskinolina maynci, corresponds to the so-called "First Palorbitolina bed" of Lafarge (1978). The marly bed seals the LO of the genus Agriopleura, otherwise known as the "Agriopleura extinction event", just antecedent to the Gerhardtia sartousiana zone (Masse & Fenerci-Masse, 2013b; Frau et al., 2018).
Unit SM III (55 m thick) is dominated by rudist beds. The predominance of Requieniidae and Monopleuridae, e.g., Debrunia favrei, D. mutabilis (Masse & Fenerci-Masse, 2018) in these beds, aligns with the Orgon fauna of Masse et al. (2020), and dates the upper part of the Gerhardtia sartousiana Zone. Unit SM III is capped by the Palorbitolina lenticularis-Heteraster oblongus guide level, formerly assigned to the lowermost Aptian (Lafarge, 1978; Cotillon et al., 1979), its lower boundary was regarded as the "late Barremian drowning" by Masse and Fenerci-Masse (2011) but the guide level was re-assigned to the latest Barremian Imerites giraudi zone (Frau et al., 2018).
Unit SM IV (65 m thick), is dominated by requieniid-bearing beds tight or chalky (physical properties). It yields the orbitolinids Orbitolinopsis buccifer, Paracoskinolina maynci and P. sunnilandensis associated with Palorbitolina lenticularis. The Caprinidae Pachytraga paradoxa (= P. lapparenti in Lafarge, 1978) recorded in the upper part of the unit, was regarded as an index of the Rustrel fauna of Masse et al. (2020). We assume that the fossil-rich beds of this unit correspond to the "Saint-Montan quarries" mentioned by Paquier (1903, 1905) (see below). The corresponding unit is, therefore, equivalent to the U2 member of Monts-de-Vaucluse (Masse & Fenerci-Masse, 2011), hence assigned to the lower part of the Martelites sarasini zone. Unit SM IV ends with a keystone-vug-bearing horizon (Lafarge, 1978), topped by an exposure surface considered equivalent to the U2/U3 boundary of the Monts-de-Vaucluse (Masse, 1976; Tendil et al., 2018).
Unit SM V (in the range of ~40 m thick) starts with 5m-thick,
thin-bedded calcarenites, followed by two main cliffed rudist beds.
In comparison to the stratigraphic succession of northern Provence, it is
determined that the Barremian/Aptian boundary is situated within these
calcarenites (Tendil et al., 2018). Consequently, the rudist-bearing beds above with Paracoskinolina arcuata Arnaud
Vanneau (in Lafarge, 1978) are attributed to the lower Aptian pro
parte. Hence, this unit bearing rudists is proposed to be stratigraphically
equivalent to the U3 Member of the Monts-de-Vaucluse and extensive areas of the
Languedoc region, and defined as the Saint-Montan Member of the regional
Urgonian Formation.
Unit SM V is topped by a drowning surface and overlain by ammonite-bearing marls. These marls, previously referred to as the "Cheloniceras marls" (Masse & Fenerci-Masse, 2011), represent a portion of the newly defined Chabert Formation of Pictet et al. (2015) and dated to the upper part of the Deshayesites forbesi zone, specifically the Roloboceras hambrovi subzone (Pictet et al., 2015; Pictet & Delanoy, 2017; Delanoy et al., 2022; Frau et al., 2023) coincident with the OAE1a Selli event (Moullade et al., 2000).
|
|
Figure 3:
Stratigraphic log of
the reference Urgonian succession of Saint-Montan (modified after Lafarge,
1978). Facies legend: 1) marls; 2) coarse grained
calcarenites; 3-4) bioclastic
coral beds; 5) caprinid rudist; 6) Agriopleura; 7) requieniid rudist;
8) early exposure surface. |
The preceding stratigraphic account leads to the following conclusions and their implications for stratigraphic correlation. The existence of two Palorbitolina-bearing marly horizons in the Saint-Montan section as proposed by Lafarge (1978), cannot be substantiated, the upper horizon is the proper one and the position of the Barremian-Aptian boundary is reappraised. The placement advocated by Clavel et al. (2014) regarding the Imerites giraudi Zone and the Barremian/Aptian boundary is equally untenable, this interpretation is in part due to the meaning of the "Imerites giraudi Zone", which refers to the former ammonite zonation of Reboulet et al. (2009), reappraised subsequently (see Reboulet et al., 2014). Moreover this ammonite zone currently applies to the Palorbitolina lenticularis guide level, recorded upsection. The early Aptian age of the SM III and SM IV, namely the Ba5 and Bd1 sequences sensu Clavel et al. (2014), is also rejected.
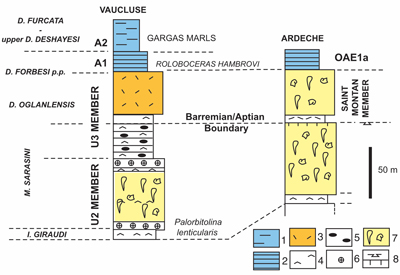
The Barremian-Aptian boundary is herein located in the basal calcarenites of the Saint-Montan-Member tentatively ascribed to the "Pseudocrioceras mazierei" beds [an index proposed by Frau and Delanoy (2022) formerly designed as Pseudocrioceras wagenoides], which coincides with the age proposed for the basal U3 Member Monts-de-Vaucluse.
The age of the marly cover aligns with that of marly cover, A1, of the U3 Member in the Monts-de-Vaucluse (Frau et al., 2018). The "Cheloniceras marls" of Masse & Fenerci-Masse (2011) atop the Urgonian limestones do not constitute a single lithostratigraphic entity. Instead, they can be categorised into two distinct lithostratigraphic units: the lower one aligning with the Chabert Formation and the upper one with the overlying Frayol Formation (sensu Pictet et al., 2015). Both formations contain Cheloniceras species but have differing ages.
To conclude, the rudist-bearing limestones forming the unit SM V of Saint-Montan, i.e., the Saint-Montan Member, formerly designed as the Bd2 unit and assigned to the "D. weissi" zone (Clavel et al., 2014) spans the Deshayesites oglanlensis and D. forbesi p.p. zones, i.e., excluding for the latter the Roloboceras hambrovi subzone (see above). Its associated rudist fauna is designated, herein, as the Saint-Montan fauna.
At Aigučze, the Saint-Montan Member is well represented in the cliffs supporting the village, its basal discontinuity being marked by a Palorbitolina lenticularis-rich horizon. In the succession figured by Bastide (2014) as "Aigučze", the Saint-Montan Member represents only the uppermost 50 m, the orbitolinids reported by the author include Orbitolinopsis pygmea and Paracoskinolina arcuata. The succession studied at the Cité du barrage correlates with the Viviers section, in which the rudist-rich Bd2 unit, ascribed to the "D. weissi" Zone (Clavel et al., 2014) is assumed to be equivalent to the Saint-Montan Member.
At Aigučze, Trescouvieux and Cité du barrage, the Saint-Montan Member and its fauna are well defined and yield a rich and diversified rudist fauna described herein.
|
|
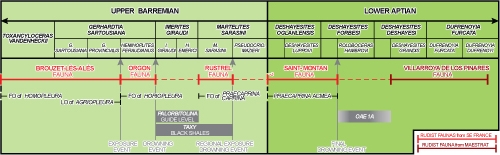
Figure 4: Stratigraphic
correlations between the upper Barremian-lower Aptian Urgonian successions of
the Monts de Vaucluse and Ardčche (modified from Masse et
al., 2020: Fig. 10). |
The present study is based
on field collections obtained from the four above-mentioned localities of early
Aptian age and the two localities from late Barremian age (see Fig. 2 ![]() ), it
includes about 200 rudist-bearing samples. The vast majority being well
lithified, they were cut with a sawing machine, and usually revealed more than
one rudist section, hence the total number of studied specimens exceeded 350.
Most were identified at family level and some of them the genus was determined.
In the following we describe the identified species of four rudist families:
Requieniidae, Monopleuridae, Caprinidae, and Polyconitidae, i.e., those
usually recorded in Urgonian rudist-bearing successions of SE France (e.g.,
Masse, 1996).
), it
includes about 200 rudist-bearing samples. The vast majority being well
lithified, they were cut with a sawing machine, and usually revealed more than
one rudist section, hence the total number of studied specimens exceeded 350.
Most were identified at family level and some of them the genus was determined.
In the following we describe the identified species of four rudist families:
Requieniidae, Monopleuridae, Caprinidae, and Polyconitidae, i.e., those
usually recorded in Urgonian rudist-bearing successions of SE France (e.g.,
Masse, 1996).
Abbreviations used for the
taxonomic descriptions and figures. LV-left valve and RV-right
valve, are used in the text.
The following are
essentially used in the figures. P-posterior, D-dorsal, V-ventral, BC-body
cavity, dap-antero-posterior diameter, dvd-dorso-ventral diameter, am-anterior
myophore, pm-posterior myophore, mb-myophoral bulge, pmt-posterior myophoral
tract, pmcr-posterior myophoral crest, amcr-anterior myophoral crest,
pc-posterior cavity, Vc-ventral carina, vr-inner ventral ridge, rb-rib,
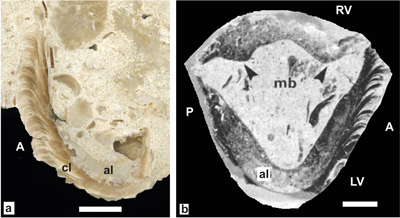
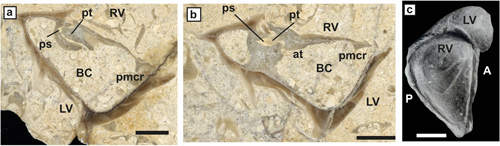
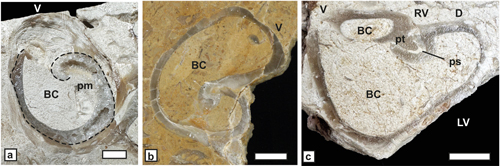
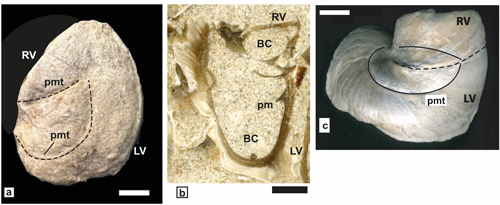
ca-carina, at-anterior tooth, pt-posterior tooth, as-anterior tooth socket, ps-posterior tooth socket, al-former aragonitic inner shell layer,
cl-outer
calcitic shell layer, AB-Anterior band, PB-posterior band, IB-interband, lg-ligament groove,
com-commissure.
Abbreviations used for the
figures of the Caprinidae refer to myophoral and perimyophoral cavities:
Ppmc-perimyophoral cavity (posterior ectomyophoral cavity of the RV in Rineau
et al., 2020), mc-myophoral cavity,
with A or P for anterior and posterior (pmc-LV is equivalent to the central
tooth socket, and pmc-RV to the posterior endomyophoral cavity in Rineau et
al., 2020).
Samples collected by C. Frau, J-P. Masse, and M. Fenerci-Masse, are housed at the Musée de Paléontologie, Aix-Marseille University, Centre Saint-Charles (specimens JPMA), other specimens collected by V. Rineau are housed in the paleontological collections of Sorbonne Université (Paris) (specimens SU.PAL).
The classification of the Hippuritida used herein refers to Skelton (2013a, 2013b).
Order Hippuritida Newell, 1965
Suborder Requieniidina Skelton, 2013a
Superfamily Requienioidea Kutassy, 1934
Family Requieniidae Kutassy, 1934
? Subfamily Matheroniinae Scott et al., 2010
Genus Matheronia Munier-Chalmas, 1873
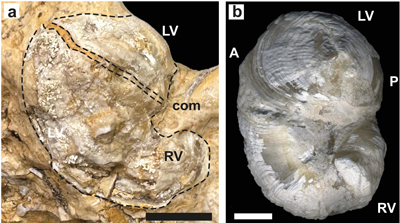
The genus Matheronia was proposed by Munier-Chalmas (1873) for Caprotina virginiae Gras (1854), the type species, but its description was due to Douvillé (1887, 1915), complemented by Paquier (1903). It is one of the oldest and most primitive members of the family Requieniidae (Masse, 2002; Skelton, 2013a). Generic characters of the genus Matheronia have been recently revised by Masse et al. (2024a) focusing on the diagnostic myophoral characters, i.e., a posterior myophoral crest and an anterior myophoral bulge on the RV, LV myophores being on the inner shell wall. By contrast Requienia Matheron and Toucasia Munier-Chalmas possess myophoral plates on RV, lacking in Matheronia, as established by Douvillé (1915). Moreover, the above revision suggests that the definition of the subfamily Matheroninae sensu Scott et al. (2010) has to be reappraised.
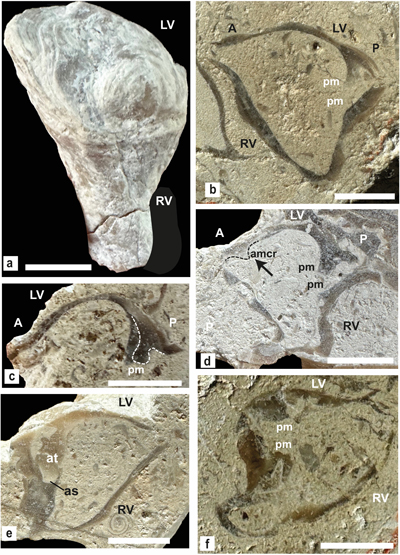
Matheronia virginiae (Gras, 1854)
(Fig. 5 ![]() )
)
The study material is
represented by a section of the anterior side of the LV (Fig.
5.a ![]() ) from Cité du
barrage (SU.PAL.2022.1.15.5.1).
) from Cité du
barrage (SU.PAL.2022.1.15.5.1).
Description. Outer shell
layer, originally calcitic, characterized by its reflexed lamellar structure,
bent outwards (Fig.
5.a ![]() ) unique in the Requieniidae, illustrated, for comparison,
by a longitudinal section of a specimen from the late Barremian Rustrel fauna of
the Monts-de-Vaucluse (Fig. 5.b
) unique in the Requieniidae, illustrated, for comparison,
by a longitudinal section of a specimen from the late Barremian Rustrel fauna of
the Monts-de-Vaucluse (Fig. 5.b ![]() ), showing the myophoral bulges (mb) (a
reproduction of Plate 5, fig. 5 in Masse, 1996). Shell thickness up to 10
mm, as in the material from the Monts-de-Vaucluse and thinner than in specimens
from the subalpine region, which can reach 20 mm in thickness (Paquier,
1903).
), showing the myophoral bulges (mb) (a
reproduction of Plate 5, fig. 5 in Masse, 1996). Shell thickness up to 10
mm, as in the material from the Monts-de-Vaucluse and thinner than in specimens
from the subalpine region, which can reach 20 mm in thickness (Paquier,
1903).
|
|
Figure 5:
Matheronia virginiae (Gras). a) Section of reflexed lamellar
anterior calcitic shell layer of LV from Cité du barrage
(SU.PAL.2022.1.15.5.1). b) Section of a bivalve specimen illustrated by Masse,
1996, from the Rustrel fauna in the Monts-de-Vaucluse. Scale bar is 10 mm. |
Age. Matheronia virginiae was recorded by Paquier (1903) at les Rimets-Vercors (type locality) and Saint-Montan. It is a common species of the late Barremian Rustrel fauna, which extends to the early Aptian (Masse et al., 2020)
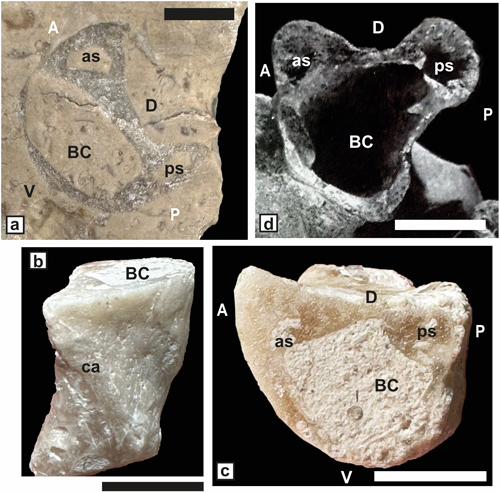
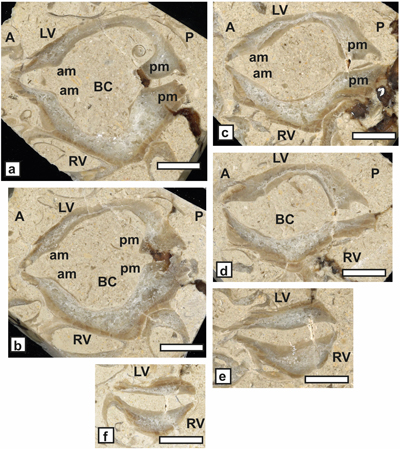
Matheronia aff. aptiensis Matheron, 1879
(Fig. 6 ![]() )
)
Study material. Two dorso-ventral sections of a single bivalve specimen (SU.PAL.2022.1.15.14.1) from Saint-Montan.
Generic placement. The "spoon like" shape (= "en
cuillčre" sensu
Masse, 2002) (Fig.
6.a-b ![]() ) of the posterior socket implanted on the inner
margin of the LV cardinal plateau is a requienid character which typifies the
cardinal apparatus of this valve. The flattened RV and the presence of a
posterior myophoral crest (pmc of Fig. 6.a-b
) of the posterior socket implanted on the inner
margin of the LV cardinal plateau is a requienid character which typifies the
cardinal apparatus of this valve. The flattened RV and the presence of a
posterior myophoral crest (pmc of Fig. 6.a-b ![]() ) typifies the genus Matheronia (Masse et
al., 2024a).
) typifies the genus Matheronia (Masse et
al., 2024a).
Description. The presence
of a vigorous ventral carina and one subordinate acute carina flanking the
dorsal side of the LV are typical morphological traits; the dorsal side of the
LV is at low angle with the RV plane, giving the longitudinal cross-sectional
habit a subtriangular shape. Sections suggest that the lamellar anterior side of
the LV is not reflexed as in M. virginiae.
The size of our specimens are smaller (Ddv = 40 mm) than that of west-European
species, M. virginiae and M. munieri,
and from the late Barremian-early Aptian Carpatho-Cimmerian species (Masse et
al., 2024a). The Barremian species, M.
aptiensis Matheron, has a comparable size, carinate habit and
a short umbonal cavity (see Paquier, 1903: Pl. III, fig. 6 reproduced herein,
Fig. 6.c ![]() ), but a more inflated RV dorsally and a limited anterior shell foliation on
LV. Our specimen is, therefore, close to those from Orgon described by Paquier
(1903).
), but a more inflated RV dorsally and a limited anterior shell foliation on
LV. Our specimen is, therefore, close to those from Orgon described by Paquier
(1903).
|
|
Figure 6: Matheronia
aff. aptiensis Matheron.
a-b) Oblique dorso-ventral pair sections of a bivalve specimen showing the
internal characters (SU.PAL.2022.1.15.14.1) from Saint-Montan. c) Matheronia
aptiensis from Orgon (reproduced from Pl. III, fig. 6 in Paquier,
1903). Scale bar 10 mm. |
Age. The original material
from Matheron (1879) is represented by internal moulds from Apt, assumed
to derive from the U2 Member of the Monts de Vaucluse (Masse, 1976),
formerly assigned to the lower Aptian, but the species is a member of the upper
Barremian Rustrel fauna (Masse et
al., 2020). Well preserved specimens from Orgon, described by Paquier
(1903, e.g., Fig. 6.c ![]() reproduced herein), belongs to the Orgon fauna.
Assuming that the identification of our specimen is correct, Matheronia
aptiensis extends to the early Aptian.
reproduced herein), belongs to the Orgon fauna.
Assuming that the identification of our specimen is correct, Matheronia
aptiensis extends to the early Aptian.
Genus Requienia Matheron, 1842
The genus with type species Chama ammonia Goldfuss, 1837, is characterized by the presence of a posterior myophoral plate on RV (Douvillé, 1915).
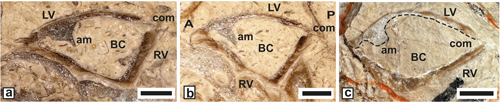
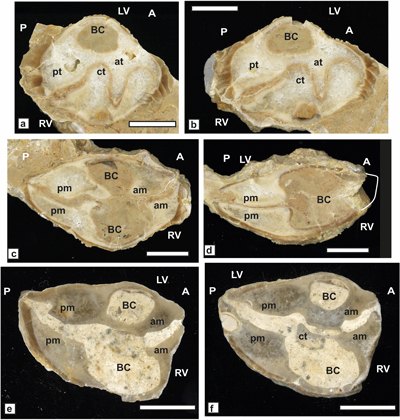
Requienia ammonia (Goldfuss, 1837)
(Fig. 7 ![]() )
)
Study material and
description. This large, thick-shelled requieniid (Dap 40-50 mm) endowed with a
posterior plate on RV, is relatively common and represented by randomly oriented
sections, our material comes from Aigučze. A section near the commissure (JPMA
18082-1A), shows the oval transverse outline (Ddv = 60 mm), the spirogyrate
growth of the posterior myophoral track and a portion of the slightly depressed
RV outer shell surface (Fig. 7.a ![]() ). A section of a LV (SU.PAL.2022.1.15.12.1)
shows the coiling habit and the convex ventral outline (Fig.
7.b
). A section of a LV (SU.PAL.2022.1.15.12.1)
shows the coiling habit and the convex ventral outline (Fig.
7.b ![]() ). A dorsal
section of bivalve specimen (JPMA 18090) shows the myophoral arrangement of the
RV and the cardinal connection of the valves (Fig. 7.c
). A dorsal
section of bivalve specimen (JPMA 18090) shows the myophoral arrangement of the
RV and the cardinal connection of the valves (Fig. 7.c ![]() ).
).
|
|
Figure 7: Requienia ammonia
(Goldfuss) from Aigučze. a) External view of a RV
showing the spirogyrate underlined track of the posterior myophore (JPMA
18082-1A). b) Dorso-ventral oblique section of a LV showing the coiling habit
and thickened, convex calcitic outer shell (SU.PAL.2022.1.15.12.1). c)
Dorso-ventral section of a bivalve specimen showing the RV myophore and the
spoon-like shape of the posterior socket of LV and salient tooth of RV (JPMA
18090). Scale bar 10 mm. |
Age. Our data show that Requienia ammonia, hitherto, documented from the Barremian (Masse et al., 2020) extends to the early Aptian.
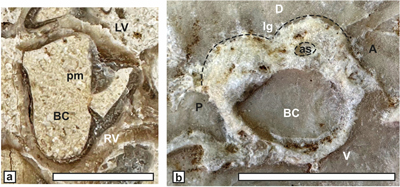
Requienia gryphoides Matheron, 1842
(Fig. 8 ![]() )
)
Material. We refer to a well-preserved LV (JPMA 18136a) from Aigučze, lacking the RV and having dimensions (Dap = 30 mm, Ddv = 30 mm, total length = 40 mm) similar to the specimens from Orgon.
Description.
This taxon originally assigned to Requienia
by Matheron (1842) was then placed in Matheronia by the same author (Matheron,
1878), an option
followed by Paquier (1903) but rejected by Douvillé (1915),
because its RV myophoral plate conforms to that of Requienia. The species
(Fig. 8 ![]() herein) has a flattened anterior side
and a rounded posterior side, with weak, poorly preserved radial bands, and it
bears a vigorous, nodulous carina (nc) at the antero-posterior junction and a
short beak.
herein) has a flattened anterior side
and a rounded posterior side, with weak, poorly preserved radial bands, and it
bears a vigorous, nodulous carina (nc) at the antero-posterior junction and a
short beak.
Age. The species is common in the late Barremian Orgon and Rustrel faunas (Masse et al., 2020) but as shown herein and from southern Spain (Masse et al., 1998) it extends to the early Aptian.
|
|
Figure 8: Requienia gryphoides Matheron from
Aigučze. View of a LV
showing the ventral nodulous carina (nc) and short beak (JPMA 18136a). Scale bar
10 mm. |
Genus Toucasia Munier-Chalmas, 1873
In this genus, with type species Requienia carinata Matheron, 1842, there is a posterior myophoral plate on both the RV and on the LV, which distinguishes it from Requienia and Matheronia, in which it is lacking in the LV (Douvillé, 1915). The distinction of species is based on the habit of the posterior shell side and the offset of the dorsal margin of the RV above the commissure (Paquier, 1903), the geometrical relationships between the LV and RV posterior myophores (Douvillé, 1918) and the morphometric W/L ratio, i.e., width/length relationships, of Masse et al. (1998), an expression of the antero-posterior shell compression. The identification of species in cross section is usually problematic.
Toucasia carinata (Matheron, 1842)
(Fig. 9 ![]() )
)
Material. Our material consists of an internal mold from Aigučze (JPMA 18067) and a section (SU.PAL.2022.1.15.5.2) from the Cité du barrage.
Description. The RV of this
species has a well-marked posterior carina and the corresponding flank is not
salient above the commissure. The ventral carina on LV is also a prominent
character of the species. On the internal mold (Fig. 9.a ![]() ) the myophores are
represented by grooves intersecting at the commissure at a relatively high angle
(50-60°), the LV myophore is quite far from the commissure, a feature
recognized by Douvillé (1918) as a criterion for T.
carinata (Fig. 9.c
) the myophores are
represented by grooves intersecting at the commissure at a relatively high angle
(50-60°), the LV myophore is quite far from the commissure, a feature
recognized by Douvillé (1918) as a criterion for T.
carinata (Fig. 9.c ![]() ). The W/L ratio = 0.4 (3/8 cm) conforms to that of T.
carinata var. compressa of Paquier (1903) a possible independent species (see
Masse, 1976). The transverse section
(Fig. 9.b
). The W/L ratio = 0.4 (3/8 cm) conforms to that of T.
carinata var. compressa of Paquier (1903) a possible independent species (see
Masse, 1976). The transverse section
(Fig. 9.b ![]() ) cuts the posterior
myophores and the thin cardinal platform, and shows the asymmetry between the
smooth posterior and lamellar anterior sides of the LV. Figure
9.c
) cuts the posterior
myophores and the thin cardinal platform, and shows the asymmetry between the
smooth posterior and lamellar anterior sides of the LV. Figure
9.c ![]() illustrates a
bivalve specimen from Orgon showing the raised dorsal margin of the RV and the
myophoral grooves (pmt) intersecting at the commissure.
illustrates a
bivalve specimen from Orgon showing the raised dorsal margin of the RV and the
myophoral grooves (pmt) intersecting at the commissure.
Age. Toucasia carinata has a wide stratigraphic extent throughout the Barremian and early Aptian (Masse et al., 2020) but we are aware that the accurate identification in the literature of the species so designated.
|
|
Figure 9: Toucasia carinata (Matheron).
a) Internal mold of a bivalve
specimen, posterior view showing the RV and LV myophoral tracks, from Aigučze (JPMA 18067).
b) Longitudinal section of a bivalve specimen showing the internal
features, from Cité du barrage (SU.PAL.2022.1.15.5.2). c) Posterior view of a
specimen from Orgon showing the posterior myophoral grooves at the shell surface
(reproduced from Masse et al., 2019). Scale bar 10 mm. |
Superfamily Radiolitoidea Orbigny, 1847
Family Monopleuridae Munier-Chalmas, 1873
Genus Monopleura Matheron, 1842
The genus is characterized by a conical RV with a posterior myophore on a transverse thickening of the anterior shell wall (Masse & Fenerci-Masse, 2020).
Monopleura varians Matheron, 1842
(Fig. 10 ![]() )
)
Study material and
description. A specimen from Aigučze (JPMA 18107). Monopleura
varians has a convex, low domal LV, a subcyndrical, short RV with a
spirogyrate apex and inconspicous radial bands (Matheron,
1842; Douvillé, 1918). The posterior myophore is tranversally thickened on both valves, a
typical attribute of the genus Monopleura and
the anterior myophore of LV as well, a typical character of this species (Masse
& Fenerci-Masse, 2020). Our bivalved specimen (Fig.
10.a ![]() ) conforms to
the morphological description given above. Its dimensions (Ddv = 20 mm, Hv = 30
mm) are in the range of those given for the specimens from Orgon figured herein
for morphological comparisons (Fig. 10.b
) conforms to
the morphological description given above. Its dimensions (Ddv = 20 mm, Hv = 30
mm) are in the range of those given for the specimens from Orgon figured herein
for morphological comparisons (Fig. 10.b ![]() ,
reproduced from Masse & Fenerci-Masse,
2020).
,
reproduced from Masse & Fenerci-Masse,
2020).
|
|
Figure 10:
Monopleura varians Matheron. a)
Posterior view of a bivalve specimen from Aigučze (JPMA 18107). b) Specimen
from Orgon (reproduced from Masse et
al., 2020: Fig. 7, H). Scale bar 10 mm. |
Age. Monopleura varians is a classical member of the Orgon fauna (Matheron, 1842) but was also recorded in the underlying Brouzet and overlying Rustrel faunas (Masse, 1976; Masse et al., 2020). As shown herein, it extends to the early Aptian.
Genus Bicornucopina Hofmann in Hofmann & Vadŕsz, 1912
The genus defined by Hofmann
(1912) and based on a single species, Bicornucopina
petersi Hofmann,
1912, the type species, was considered close to
Praecaprina and Pachytraga by Czabalay (1968) or even Caprinula by Perkins and Coogan (in Dechaseaux et
al., 1969) but placed in the group "Family uncertain" by the last
authors. The existence of "radial canals" mentioned by the former authors
and suggesting a possible affinity with the Caprinidae, has been rejected by Masse
(1992), who has provided evidence that the so-called canals correspond to radial
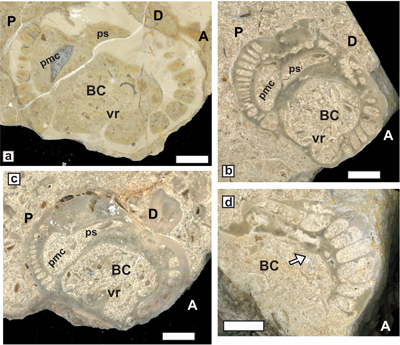
partitions developed in the two RV teeth sockets (Fig. 11.d ![]() ).
).
Bicornucopina belongs to the
Monopleuridae (Skelton, 2013a). The genus is characterized by a flat or
depressed dorsal side and convex, rounded ventral side, lacking radial bands.
Shell valves are hemi-cylindrical and arcuate. The teeth on LV are arcuate and
projecting. On RV the teeth sockets are distant and located on two salient
outward, wing-like expansions. This organization, well expressed in the type
species (Fig. 11.d ![]() ) is regarded herein as a diagnostic, generic trait. Evidence
for a bimineralic shell is lacking, and the test is assumed to have been mainly
aragonitic, but the absence of an outer calcitic shell layer may also result
from an early (?) diagenesis. The LV lamellar myophores belong to the
petalodontid type (Masse, 1992; Masse & Fenerci-Masse,
2018), but the central tooth and socket remain poorly defined. The nature and
importance of the internal radial socket partitioning and its corollary, the
longitudinally fluted teeth, combined with the morphology of the dorsal area,
are considered specific characters. The gross overall shell morphology is
somewhat similar to that of Oedomyophorus
shaybahensis Skelton, 2004, but
their dorso-ventral shell asymmetry and cardinal organization are different.
) is regarded herein as a diagnostic, generic trait. Evidence
for a bimineralic shell is lacking, and the test is assumed to have been mainly
aragonitic, but the absence of an outer calcitic shell layer may also result
from an early (?) diagenesis. The LV lamellar myophores belong to the
petalodontid type (Masse, 1992; Masse & Fenerci-Masse,
2018), but the central tooth and socket remain poorly defined. The nature and
importance of the internal radial socket partitioning and its corollary, the
longitudinally fluted teeth, combined with the morphology of the dorsal area,
are considered specific characters. The gross overall shell morphology is
somewhat similar to that of Oedomyophorus
shaybahensis Skelton, 2004, but
their dorso-ventral shell asymmetry and cardinal organization are different.
Bicornucopina acuminata n. sp.
(Fig. 11 ![]() )
)
Fossil material. The studied material includes a single slab from Saint-Montan (JPMA 18061-13b), and two RV transversally cut (JPMA 18093a, b) from Chaux de la Tour quarry.
Holotype. RV shell section JPMA 18093a, from Chaux de la Tour quarry.
Generic attribution. The overall sparitic structure shows that the shell seems to have been essentially aragonitic. Transverse sections of the RV display a depressed dorsal side and a convex, rounded ventral side lacking radial bands; teeth sockets are located at the tip of wing-like expansions. The RV has an arcuate twisted habit, but data on the overall shell morphology and myophoral organization are not quite clear, and it is worth noting that the myophoral characters of B. petersi are also poorly documented.
Derivation of name. The two salient wing-like expansions are acuminate.
Diagnosis.
Bicornucopina with small and
subrounded tooth socket and acuminate antero-posterior cardinal wing expansions
on RV (Fig. 11.a-c ![]() ).
).
Description
of the new species and comparisons. Assuming that Bicornucopina acuminata
belongs to the genus in question, noticeable differences exist between the
Urgonian species from SE France and its Hungarian and Italian counterparts (see Masse,
1992), mainly the size and shape of the RV tooth sockets: small and subrounded
instead of large and densely, radially partitioned, and the acuminate
termination of the cardinal wing expansions of RV. The geometry of the sockets (Fig.
11.a ![]() ) suggests that the teeth were probably rather smooth, i.e.,
not fluted like the Hungarian and Italian taxa, and bifid. The ventral side is
thinner than the dorsal one, a modest radial thickening of the assumed posterior
side (flanking the assumed posterior socket) may be a myophoral trait, the
anterior side is more expanded, convex outside, than the flattened posteriorly.
The anterior side is associated with carina (Fig. 11.b-c
) suggests that the teeth were probably rather smooth, i.e.,
not fluted like the Hungarian and Italian taxa, and bifid. The ventral side is
thinner than the dorsal one, a modest radial thickening of the assumed posterior
side (flanking the assumed posterior socket) may be a myophoral trait, the
anterior side is more expanded, convex outside, than the flattened posteriorly.
The anterior side is associated with carina (Fig. 11.b-c ![]() ) and the socket is
larger than the posterior one. The size, referred to Dap (tip to tip distance)
is 15-20 mm, and Ddv 12-13 mm, comparable to that of the Hungarian type material
and Italian forms. Notice that the size of lower Aptian specimen from Saint-Montan (Fig. 11.a
) and the socket is
larger than the posterior one. The size, referred to Dap (tip to tip distance)
is 15-20 mm, and Ddv 12-13 mm, comparable to that of the Hungarian type material
and Italian forms. Notice that the size of lower Aptian specimen from Saint-Montan (Fig. 11.a ![]() ) is larger (Dap = 25 mm, Ddv = 15 mm) than that of the
upper Barremian, and the size of the sockets as well.
) is larger (Dap = 25 mm, Ddv = 15 mm) than that of the
upper Barremian, and the size of the sockets as well.
Environmental, palaeoecological settings and age. In its two localities of SE France Bicornucopina acuminata n. sp. was recorded in Urgonian polytaxic rudist assemblages including Pachytraga paradoxa, Praecaprina varians, Horiopleura brevis, and large requieniids. This mode of assemblage is similar to that of Bicornucopina cf. petersi from the lower Aptian of Italy (Masse, 1992). It contrasts with the mode of occurrence of B. petersi from Hungary, recorded in a submarine volcano-sedimentary complex forming an atoll-rimmed structure (Czabalay, 1968; Csŕszŕr & Turnšek, 1996) and bearing Valanginian foraminifera and rare rudists recorded from the Mecsek Mountains. The age is similar to that of other neighboring submarine volcanic spots (Csŕszŕr et al., 2013) but the age of the reworked associated rudists might be different, and possibly younger (Csŕszŕr, personal communication).
Age. The age of the type species is uncertain, B. cf. petersi is an early Aptian species. B. acuminata n. sp. is recorded in the late Barremian Rustrel fauna and in the early Aptian Saint-Montan fauna.
|
|
Figure 11:
Bicornucopina acuminata
n. sp. a) Transverse section of a RV from Saint-Montan (JPMA 18061-13b), and two
RV transversally cut (JPMA 18093a, b) from Chaux de la Tour quarry). b) Chaux de
la Tour, holotype, RV showing the anterior carina (ca) (JPMA 18093a). c) Ibidem transverse view showing
the acuminate wings. d) Transverse section of the RV of Bicornucopina petersi with protruding rounded, partitioned cardinal
zones (type from Hungary in Czabalay, 1968). Scale bar 10 mm. |
Genus Myostyla n. gen.
Type species. Myostyla ardescensis n. sp.
Diagnosis. Low, shell with well-defined outer calcitic and inner formerly aragonitic layers, RV conical, LV convex, trend towards biconvexity, anterior LV myophoral plate, with a sub-triangular transverse outline, oblique to the commissure and protruding into the opposite valve, LV posterior myophore on shell wall, anterior RV thickening facing the anterior LV plate, posterior RV myophore on shell wall or on downward sloping ledge.
Taxonomic position. The shell morphology and myophoral organization suggest a placement in the family Monopleuridae with some similarities with the Polyconitidae. Referring to some members of the Monopleuridae, e.g., Monopleura (see Masse & Fenerci-Masse, 2020) or the Polyconitidae (see Masse & Fenerci-Masse, 2017) having a contrasting development of the LV anterior and posterior myophores, we may assume, by analogy, that the myophoral plate of our new genus might be posterior. Nevertheless the location of the myophoral plate on the same side as the RV shell attachment surface, typically anterior (see Skelton, 1976), suggests that the plate is more probably anterior. The presence of a single visible tooth attached to the myophoral plate, low conical and strongly developed, suggests it is the anterior tooth, generally more developed than the posterior tooth, which also supports the hypothesis that the myophoral shelf adjoining the tooth is also anterior. The morphology of the myocardinal apparatus and the sub-triangular, lamellar transverse outline warrant a placement of Myostyla n. gen. in the Monopleuridae rather than in the Polyconitidae.
Derivation of name. From the pointed termination (Latin stylos) in section, of the LV myophoral plate.
Age. Early Aptian.
Type locality. Saint-Montan.
Myostyla ardescensis n. sp.
(Fig. 12 ![]() )
)
Fossil material. Two slabs cut from Saint-Montan in a tight block
(JPMA
18061 a, b) including 2 transverse sections of a specimen with conjoined valves,
the larger (Fig. 12.a ![]() )
being the holotype for both the genus and the species. An
additional section (JPMA 18061-10) from the same locality is represented (Fig.
12.c
)
being the holotype for both the genus and the species. An
additional section (JPMA 18061-10) from the same locality is represented (Fig.
12.c ![]() ).
).
Derivation of name. From the Latin name Ardesca of Ardčche river.
Diagnosis. Myostyla with moderate
cross sectional dimensions, calcitic outer shell layer thick and smooth, shell
morphology trend towards biconvexity (Fig. 12.a-c ![]() ).
).
Description. Smooth shell, RV subconical, LV convex, trend towards
biconvexity, somewhat compressed orthogonally to the commissure, moderate size,
cross sectional dimensions: Dap = 40 mm, Ddv = 17 mm for the larger specimen. RV
thicker than LV (Fig. 12 ![]() ). The RV outer calcitic shell layer up to 5 mm thick in
the myophoral zone, then reduced to 1-2 mm; with a similar thickness on LV. The
internal formerly aragonitic shell layer 1 to 2 mm thick on both valves, it
thickens gradually on LV towards the salient myophore and on the opposite side
of RV. The LV myophore has a main axis oblique to the commissure, a marginal
insertion, connected with the anterior tooth with a triangular cross-sectional
elongated non-pedicular shape and protrudes into the RV (Fig.
12.a-b
). The RV outer calcitic shell layer up to 5 mm thick in
the myophoral zone, then reduced to 1-2 mm; with a similar thickness on LV. The
internal formerly aragonitic shell layer 1 to 2 mm thick on both valves, it
thickens gradually on LV towards the salient myophore and on the opposite side
of RV. The LV myophore has a main axis oblique to the commissure, a marginal
insertion, connected with the anterior tooth with a triangular cross-sectional
elongated non-pedicular shape and protrudes into the RV (Fig.
12.a-b ![]() ). The
significance of the thickened basal part of the myophore is unclear so
additional data on this character are needed, expected to improve the taxonomy
of this form.
). The
significance of the thickened basal part of the myophore is unclear so
additional data on this character are needed, expected to improve the taxonomy
of this form.
|
|
Figure 12:
Myostyla ardescensis n. gen. and n. sp. from Saint-Montan. a-b)
Conjugate antero-posterior sections (JPMA 18061a, b) showing the cross-sectional
triangular shape of the LV lamellar myophore, in (a) the inflated basal portion
may represent the teeth-myophore junction. c) Ibidem, showing the
teeth-myophore
connection. Scale bar 10 mm. |
Genus Debrunia Masse & Fenerci-Masse, 2009
The genus Debrunia Masse & Fenerci-Masse (2009), with type species Monopleura mutabilis Matheron (1878), is a cylindro-conical monopleurid characterized by asymmetric, erect myophoral apophyses in LV, the posterior is a plate and the anterior a crest, RV myophores are on shell wall (Masse & Fenerci-Masse, 2018).
Debrunia occitanica Masse & Fenerci-Masse, 2010
(Fig. 13 ![]() )
)
Study material and description. Longitudinal section of a specimen with two valves (JPMA 18177-2) from Aigučze, and a transverse section from Saint-Montan (JPMA 18062a).
In D. occitanica the LV is a low dome and the posterior myophore well
developed. In the figured longitudinal section (Fig. 13.a ![]() ) the posterior
myophore on RV is a ledge, a feature not documented before in this species and
even in other species of Debrunia. The
average diameter is in the range of 1-1.2 cm. the transverse section from Saint-Montan
(Fig. 13.b
) the posterior
myophore on RV is a ledge, a feature not documented before in this species and
even in other species of Debrunia. The
average diameter is in the range of 1-1.2 cm. the transverse section from Saint-Montan
(Fig. 13.b ![]() ) has a rather smooth shell, inconspicuous radial bands
and the bilobate dorsal margin, an attribute of the species (Masse & Fenerci-Masse,
2010).
) has a rather smooth shell, inconspicuous radial bands
and the bilobate dorsal margin, an attribute of the species (Masse & Fenerci-Masse,
2010).
Age. Debrunia occitanica was documented from the late Barremian Orgon and Rustrel faunas (Masse et al., 2020), its record at Aigučze and Saint-Montan shows that it also occurs in the early Aptian.
|
|
Figure 13:
Debrunia occitanica Masse & Fenerci-Masse. a)
Longitudinal section of a bivalve specimen showing the posterior myophoral organization, from Aigučze
(JPMA 18177-2). b) Transverse section of a RV showing
the bilobate dorsal side, from Saint-Montan (JPMA 18062a). Scale bar 10 mm. |
Debrunia ucetiae Masse & Fenerci-Masse, 2018
(Fig. 14 ![]() )
)
Study material and
description. A slab (JPMA 18061-2) from Saint-Montan cuts a shell aggregation
including numerous individuals grouped in a vertical bundle (Fig.
14.a ![]() ). Shells
are essentially represented by RV with diameters in the range of 1-1.5 cm,
similar to the size of specimens from the type locality at Uzčs. They show the
longitudinal, acute ribs cut vertically or transversally, which characterize the
species, the type figure of which is given (Fig. 14.b
). Shells
are essentially represented by RV with diameters in the range of 1-1.5 cm,
similar to the size of specimens from the type locality at Uzčs. They show the
longitudinal, acute ribs cut vertically or transversally, which characterize the
species, the type figure of which is given (Fig. 14.b ![]() ) herein for comparison and
reproduced from Figure 14.D in Masse and Fenerci-Masse
(2018).
) herein for comparison and
reproduced from Figure 14.D in Masse and Fenerci-Masse
(2018).
Age. Debrunia ucetiae is a member of the Orgon and Rustrel faunas of late Barremian age; the present record documents its occurence in the lower Aptian rock units.
|
|
Figure 14: Debrunia ucetiae Masse & Fenerci-Masse.
a) Section of a monospecific shell bundle
(ribbed RV dominated) from Saint-Montan. b) Type specimen from
Uzčs. Scale
bar 10 mm. |
Genus Mathesia (Mainelli, 1996)
The morphological and myocardinal characters of the genus Mathesia, with its type species, Agria darderi Astre (1933), are similar to that of Debrunia Masse & Fenerci-Masse, but the wall architecture of the RV is different and consists of scalloped, festooned, tubular or vermiform microstructures located at the inner face of the calcitic outer shell layer (Fenerci-Masse et al., 2011).
Mathesia darderi (Astre, 1933)
(Fig. 15 ![]() )
)
Study material and
description. The studied specimens are represented by five transverse sections
of RV preserved on a rock surface (SU.PAL.2022.1.15.9.1) from la Combe des Pins,
and one in thin section (SU.PAL. 2022.1.15.12.4) from Aigučze (Fig.
15.a ![]() ). The
macroscopic specimens show a calcitic outer shell layer with short acute ribs.
The ventral portion bears two concave depressions interpreted as radial bands,
some are bounded by ribs including the ventral rib (Vc). The inner part of the
calcitic shell layer is characterized by a tubular microstructure
macroscopically having a striated habit (Masse & Fenerci-Masse,
2010; Fenerci-Masse et al.,
2011). The thin section from a RV (Fig. 15.b
). The
macroscopic specimens show a calcitic outer shell layer with short acute ribs.
The ventral portion bears two concave depressions interpreted as radial bands,
some are bounded by ribs including the ventral rib (Vc). The inner part of the
calcitic shell layer is characterized by a tubular microstructure
macroscopically having a striated habit (Masse & Fenerci-Masse,
2010; Fenerci-Masse et al.,
2011). The thin section from a RV (Fig. 15.b ![]() ) shows transverse sections of a raw
of tubes and associated inward-protruding fibrous structures of the outer
calcitic shell layer, the inner formerly aragonitic shell layer is poorly
defined. The tubular structure and associated fibrous microstructure conform to
that of Mathesia darderi the organization of which is somewhat variable, the
invagination of the festooned inner shell surface may produce the segregation of
the marginal festoons as isolated tubes within the shell layer (see figures in Fenerci-Masse
et al. 2011).
) shows transverse sections of a raw
of tubes and associated inward-protruding fibrous structures of the outer
calcitic shell layer, the inner formerly aragonitic shell layer is poorly
defined. The tubular structure and associated fibrous microstructure conform to
that of Mathesia darderi the organization of which is somewhat variable, the
invagination of the festooned inner shell surface may produce the segregation of
the marginal festoons as isolated tubes within the shell layer (see figures in Fenerci-Masse
et al. 2011).
|
|
Figure 15:
Mathesia
darderi (Astre). a) Combe des Pins - upper Barremian (SU.PAL.2022.1.15.9.1).
Cluster of RV showing the transverse circular shell habit and the variable
aspect of the concave radial bands bounded by ribs. Natural section. Arrows
point to the tubular structure of the inner margin of the outer calcitic shell
layer. Scale bar 10 mm. b) Aigučze-lower Aptian (Specimen SU.PAL.2022.1.15.12.4). Thin section from a RV showing transverse sections of a raw of
tubes and associated inward protruding fibrous structures of the outer calcitic
shell layer, inner shell layer poorly defined. Arrrow Points to the inner shell.
Scale bar 0.5 mm. |
Age. As shown earlier, the stratigraphic history of the species is marked by a change in size. The maximum diameters of our specimens are in the range of 14-15 mm, a value matching that of late Barremian-early Aptian forms (Fenerci-Masse et al., 2011). The cluster of specimens of Mathesia darderi described herein is assigned to the M. sarasini zone. Its occurrence is slightly younger than the Bulgarian specimens known so far as the oldest ones and placed in the G. sartousiana zone (Fenerci-Masse et al., 2011). Specimens from Aigučze are early Aptian. Prior to its late Aptian-Albian expansion, Mathesia was, until now, essentially documented from Spain and Bulgaria (Fenerci-Masse et al., 2011). Early Aptian forms were hitherto recorded from Spain, the specimens from Aigučze represent the first mention of the species of this age in SE France, and the same is true for the Barremian occurrence from Combe des Pins. Barremian occurences of the taxon are not rare, accordingly the taxon was figured (but not formally identified) by Stein et al. (2012: Fig. 4.F) from the Rawil Member of the Urgonian limestones of the Helvetic region (Switzerland) currently assigned to the upper Barremian.
Genus Valclusella n. gen.
Type species. Valclusella valclusensis n. sp.
Derivation of name. From its presence near the village of Fontaine-de-Vaucluse, val clusa, the Latin name for "closed valley", home of the corresponding famous spring.
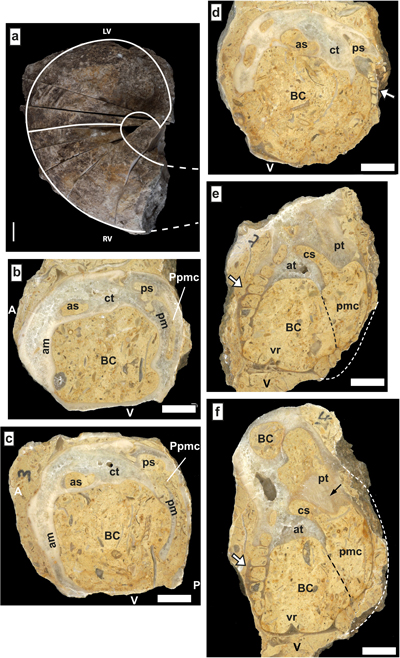
Diagnosis. RV smooth, subconical, with dorso-ventral asymmetry, on dorso-ventral longitudinal sections the dorsal side is concave and the ventral side convex; antero-posterior longitudinal sections also show an antero-posterior asymmetry, the anterior side more expanded and slightly convex than the posterior side, shorter and slightly concave. LV smooth to lamellar, domal, with a low convexity, salient rounded beak prosogyrate or opisthogyrate, interrupted at the dorsal commissural edge. Radial bands poorly defined, ventral side with wavy growth lines. LV posterior myophore vertical, straight, inwardly directed and protruding in the RV. Presence of a minute posterior myophoral depression on well developed posterior accessory cavity. Anterior myophore highly variable on the LV shell wall, represented by a short vertical crest alongside an anterior accessory cavity.
Discussion. The LV posterior myophore is not reflexed or arcuate, the myophoral attachment surface is apical and not parietal as in Debrunia. The myophore does not issue from an internal, dorsal, conical cavity like in the Polyconitidae. In this Family the LV posterior myophore is a bent, twisted lamina consisting of a plate subparalell to the commissure and attached basically to the shell by a pedicle (Skelton et al., 2010; Rineau et al., 2020; Masse & Fenerci-Masse, 2021). Hence, Valclusella n. gen. is assigned to the Monopleuridae and not to the Polyconitidae. Similarities with other Monopleuridae include the overall shell morphology; conical RV and "capuloid" LV.
The distinct species belonging to Valclusella n. gen. show different traits of the diagnostic LV posterior myophore, especially regarding its size and length. The LV anterior myophore varies according to species. It is a crest in V. valclusensis n. sp. whereas it is on the shell wall in V. biconvexa n. sp. RV myophores tend to be on the shell wall, the posterior being on a shell transverse or oblique thickening, or on a thin ledge.
Valclusella valclusensis n. sp.
(Fig. 16 ![]() )
)
Derivation of name. From the type locality of the genus.
Fossil material. One isolated bivalve specimen (holotype, JPMA 18093-1), four sections of bivalve specimens, two sections and an internal mold of an isolated LV, paratypes numbers JPMA 18093-3, 4, 5 to 10 and 18065-2; all from the type locality. One from the Saint-Montan Member.
Diagnosis. Valclusella with conical RV, low domal LV, LV anterior myophore a short crest.
Age. Uppermost Barremian (M. sarasini Zone) and lower Aptian.
Type locality. Les Chaux de la Tour quarry (Robion), near Fontaine-de-Vaucluse.
Specific traits include the
characters of the genus, i.e., the thick, distally squared LV posterior
myophore on a vertical blade alongside the posterior accessory cavity (Fig.
16.b-d ![]() ), the short anterior crest (Fig. 16.d
), the short anterior crest (Fig. 16.d ![]() ) being a specific character. The
morphology (Fig. 16.a-c
) being a specific character. The
morphology (Fig. 16.a-c ![]() ) is characterized by a modest shell size with Dap barely
exceeding Dvd values. Based on sections and a single bivalve specimen, the
holotype, size
values based on sections must be considered with caution, dimensions of the
holotype, Dap = 32 mm, Dvd = 28 mm, Vh = 37 mm, Dh = 27 mm, much higher, been
more reliable. The average and extreme size measurements of specimens are as
follows: Dap = 23 mm (20-26 mm), Ddv = 25 mm (20-28 mm), H = 26 mm (25-28 mm),
HLV = 6.5 mm (6-10 mm). The dorsal side is straight or concave and is shorter
than the convex ventral side (Fig. 16.e-f
) is characterized by a modest shell size with Dap barely
exceeding Dvd values. Based on sections and a single bivalve specimen, the
holotype, size
values based on sections must be considered with caution, dimensions of the
holotype, Dap = 32 mm, Dvd = 28 mm, Vh = 37 mm, Dh = 27 mm, much higher, been
more reliable. The average and extreme size measurements of specimens are as
follows: Dap = 23 mm (20-26 mm), Ddv = 25 mm (20-28 mm), H = 26 mm (25-28 mm),
HLV = 6.5 mm (6-10 mm). The dorsal side is straight or concave and is shorter
than the convex ventral side (Fig. 16.e-f ![]() ). Shell thickness thin, calcitic
outer shell layer and internal formerly aragonitic shell layer in the range of
1mm each, the later up to 2 mm in the myophoral zones. Variability includes the
size and the shape of the anterior and posterior sides of the RV, especially for
attached specimens with anterior side fitting the shape of its substrate.
External traits are somewhat comparable to that of the coeval Horiopleura
brevis, but the adult size is smaller and internal characters are different.
). Shell thickness thin, calcitic
outer shell layer and internal formerly aragonitic shell layer in the range of
1mm each, the later up to 2 mm in the myophoral zones. Variability includes the
size and the shape of the anterior and posterior sides of the RV, especially for
attached specimens with anterior side fitting the shape of its substrate.
External traits are somewhat comparable to that of the coeval Horiopleura
brevis, but the adult size is smaller and internal characters are different.
|
|
Figure 16: Valclusella valclusensis n. sp. Upper Barremian, Les Chaux de la
Tour quarry, Robion. a) Antero-dorsal view of the holotype (JPMA 18093-1).
b)
Longitudinal section showing the myophoral organization. c) Antero-posterior
section of an isolated LV showing the distally squared left myophore. d) Ibidem,
the arrow points to the anterior small crest on LV (bivalve specimen. e)
Dorso-ventral section showing the anterior cardinal area).
f) Ibidem, showing
the concave dorsal side and convex ventral side of right valve, low left valve
and remnants of myocardinal elements. Scale bar 10 mm. |
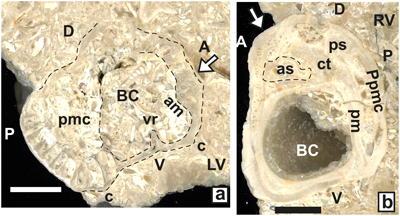
Valclusella biconvexa n. sp.
(Fig. 17 ![]() )
)
Study material. The holotype is a set of four serial sections of a single specimen from Balcon des Templiers (SU.PAL.2022.1.15.2.1). A section from Saint-Montan (JPMA 18061-12), and two sections from Aigučze (JPMA 18082 B1) are paratypes.
Diagnosis. Biconvex Valclusella with transversally moderately thickened anterior myophores, distally thickened posterior LV myophore.
Description. LV low dome,
RV inverted conical, flared, trending towards biconvexity in antero-posterior
sections. The mode of dorso-ventral decrease in size of antero-posterior serial
sections (Fig. 17.b ![]() ) documents some
dorso-ventral shell elongation, coupled with
a flattening of the RV shell (Fig. 17
) documents some
dorso-ventral shell elongation, coupled with
a flattening of the RV shell (Fig. 17 ![]() ).
Antero-posterior shell asymmetry, the
anterior side larger than the posterior one. The shell is bimineralic the
calcitic outer shell layer being thicker on RV. The LV posterior myophore
vertical alongside the posterior accessory cavity on a blade protruding in the
RV has a moderately salient squared termination and is flanked by a commarginal
furrow (Fig. 17.b-d
).
Antero-posterior shell asymmetry, the
anterior side larger than the posterior one. The shell is bimineralic the
calcitic outer shell layer being thicker on RV. The LV posterior myophore
vertical alongside the posterior accessory cavity on a blade protruding in the
RV has a moderately salient squared termination and is flanked by a commarginal
furrow (Fig. 17.b-d ![]() ). The LV myophore is flattened and parallel to the flat RV
myophoral transverse thickening, the square termination may turn to slightly
convex, the RV counterpart becoming slightly concave upward (Fig.
17.a
). The LV myophore is flattened and parallel to the flat RV
myophoral transverse thickening, the square termination may turn to slightly
convex, the RV counterpart becoming slightly concave upward (Fig.
17.a ![]() ). The LV
anterior myophore is flat or slightly concave downward. The RV anterior myophore
is on shell wall (sloping inward) or on a plateau (Fig. 17.a-d
). The LV
anterior myophore is flat or slightly concave downward. The RV anterior myophore
is on shell wall (sloping inward) or on a plateau (Fig. 17.a-d ![]() ). The shape of
the shell and the organization of the RV myophores of V.
biconvexa n. sp. strongly depart from those of V.
valclusensis and the absence of a LV anterior crest as well.
). The shape of
the shell and the organization of the RV myophores of V.
biconvexa n. sp. strongly depart from those of V.
valclusensis and the absence of a LV anterior crest as well.
|
|
Figure 17: Valclusella biconvexa n. sp. a-f) Set of serial
antero-posterior
sections of conjoined valves of the holotype from Balcon des Templiers. The
slabs show the distal reduction of LV myophores coupled with LV flattening and
progressive decrease in size (SU.PAL.2022.1.15.2.1). Scale bar 10 mm. |
Age. At Balcon des Templiers our specimens are associated with the Rustrel fauna, assigned to the upper Barremian (M. sarasini Zone). The specimens from Saint-Montan, Aigučze and Cité du barrage are lower Aptian. The presence of V. biconvexa n. sp. at three distinct lower Aptian sites shows that the species is probably rather common in the corresponding beds. Its record in the Rustrel and possibly Brouzet faunas, testifies an upper Barremian-lower Aptian stratigraphic extent.
Genus Homopleura Masse & Fenerci-Masse, 2019
The genus Homopleura Masse & Fenerci-Masse (2019), with type species Monopleura affinis Matheron (1878), differs from Monopleura in having RV anterior and posterior transverse myophoral thickenings.
Homopleura parva n. sp.
(Fig. 18 ![]() )
)
Material. Two bivalve specimens from Trescouvieux (SU.PAL.2022.1.15.11.1) and Aigučze (SU.PAL.2022.1.15.12.3), cut longitudinally in two and three slabs respectively.
Taxonomic position. The placement of our specimens in Homopleura is based on the presence of RV anterior and posterior transverse myophoral thickenings subparallel to the commissural plane.
Derivation of name. From its small size, parvus in Latin.
Holotype. Specimen from
Aigučze (Fig. 18.a-d ![]() ).
).
Diagnosis. Small globular Homopleura, LV myophores convex downward, RV myophores concave upward.
Description. Short, sub-globular
form, asymmetric with a posterior side much larger than the anterior and salient
dorsal side. Outer shell layer calcitic brownish, thin, mostly preserved on RV,
inner shell layer pale beige or translucent sparry mosaic. Shell height smaller
(22-25 mm) than commissural diameter (32-34 mm) except on dorsal sections (Fig.
18.c ![]() ). Cup like RV with an anterior outer groove or depression, posterior and
anterior myophores sloping inward, slightly depressed, the posterior larger than
the anterior, central tooth small. LV low, domal, with myophores convex downward,
the posterior larger is slightly protruding, creating posteriorly a small
posterior accessory cavity (Fig. 18.a
). Cup like RV with an anterior outer groove or depression, posterior and
anterior myophores sloping inward, slightly depressed, the posterior larger than
the anterior, central tooth small. LV low, domal, with myophores convex downward,
the posterior larger is slightly protruding, creating posteriorly a small
posterior accessory cavity (Fig. 18.a ![]() ), body cavity modest (dimension less than
half of the total diameter). Radial bands are inconspicuous.
), body cavity modest (dimension less than
half of the total diameter). Radial bands are inconspicuous.
The inconspicuous radial bands and salient LV posterior myophore are somewhat similar to those of Homopleura balkanica Masse & Fenerci-Masse, 2019, but the size is much smaller, the shape more rounded and the antero-posterior shell asymmetry is reverse.
|
|
Figure 18: Homopleura parva n. sp.
a-d)
Antero-posterior conjugate sections showing from dorsal to ventral
showing the shell outline and the internal myocardinal organization, from Aigučze
(SU.PAL.2022.1.15.12.3). e-f) Ibidem, sections from Trescouvieux
(SU.PAL.2022.1.15.11.1). Scale bar 10 mm. |
Age. Homopleura parva n. sp. is the youngest representative of the genus in Western Europe and, so far, the only well documented early Aptian species.
Family Caprinidae Orbigny, 1847
Genus Pachytraga Paquier, 1900
Type species. Sphaerulites paradoxa Pictet & Campiche, 1869
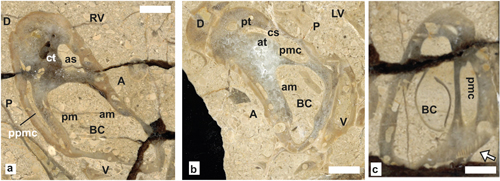
The genus is characterized by a posterior myophoral plate on RV projecting into a myophoral cavity on LV: i.e., caprinid mode of Mac Gillavry (1937; Skelton & Masse, 1998). The RV myophore is flanked by a posterior (perimyophoral) cavity. A narrow anterior myophoral cavity is on LV. Pallial canals are variously developed, depending on species.
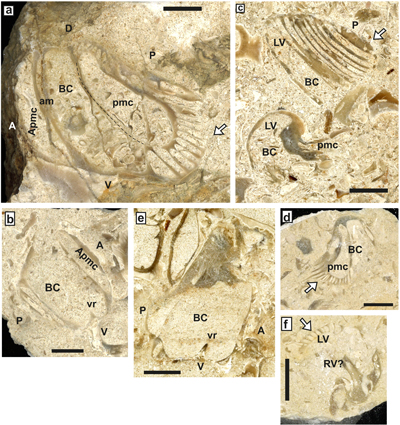
Pachytraga paradoxa (Pictet & Campiche, 1869)
Material. Recorded at la Combe des Pins (La Madeleine), P. paradoxa is illustrated by three sections, two LV and one RV (SU.PAL.2022.1.15.8.1), and paired sections of a recumbent bivalve specimen (SU.PAL.2022.1.15.8.2).
Description. The main
characters of the species outlined by Paquier (1905) and Skelton
and Masse (1998), are a cross sectional subtrapezoidal or triangular
shell outline, rounded dorsal side, rounded antero-ventral carina, posterior
side slightly convex, anterior side flat and elongated, ventral side wide, flat
to depressed. The RV (Fig. 19.a ![]() ) bears a narrow and elongated perimyophoral
cavity flanking the posterior myophore (pm), the anterior myophore (am) is
shorter and inwardly salient. Anterior socket wide and larger than the posterior
and presence of a low mid-ventral inner ridge on both valves. The LV (Fig.
19.b
) bears a narrow and elongated perimyophoral
cavity flanking the posterior myophore (pm), the anterior myophore (am) is
shorter and inwardly salient. Anterior socket wide and larger than the posterior
and presence of a low mid-ventral inner ridge on both valves. The LV (Fig.
19.b ![]() )
has a well-defined posterior myophoral cavity (pmc), the perimyophoral anterior
cavity is modest and may possess transverse partitions. The row of rectangular
canals extending on the RV anterior margin considered by Chartrousse
(1998) as a diagnostic character of the species, is missing in many specimens.
The presence of a row of rectangular canals on the anterior side of the LV is
also facultative (see Skelton & Masse,
1998). Moreover, our
material shows that canals may exist on the posterior side of the LV of some
specimens (298001b) (Fig. 19.c
)
has a well-defined posterior myophoral cavity (pmc), the perimyophoral anterior
cavity is modest and may possess transverse partitions. The row of rectangular
canals extending on the RV anterior margin considered by Chartrousse
(1998) as a diagnostic character of the species, is missing in many specimens.
The presence of a row of rectangular canals on the anterior side of the LV is
also facultative (see Skelton & Masse,
1998). Moreover, our
material shows that canals may exist on the posterior side of the LV of some
specimens (298001b) (Fig. 19.c ![]() ). Variability is, therefore, a remarkable
character of the species, already noticed by Skelton and Masse
(1998).
). Variability is, therefore, a remarkable
character of the species, already noticed by Skelton and Masse
(1998).
|
|
Figure 19: Pachytraga paradoxa (Pictet & Campiche) Combe des
Pins. a-b) Sections of RV (a) and LV (b) showing the internal characters.
c)
Section of a LV with canals developed on the postero-ventral side adjacent to
the posterior myophoral cavity (Arrow) (SU.PAL.2022.1.15.8.1). Scale bar 10 mm. |
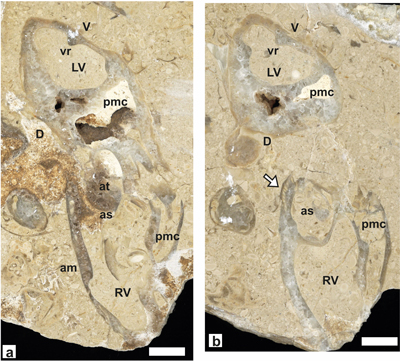
Paired sections of a
recumbent bivalve specimen are cut conjoined (a) and disjoined (b) RV and LV (Fig. 20 ![]() ). Cross-section of LV is subtriangular with mid-ventral inner ridge
(a-b), RV (a) with anterior myophoral thickening (with small rounded canals).
The anterior tooth of LV projects into the anterior socket of RV (a); the RV has
a wide perimyophoral cavity and a broken (a) and a preserved (b) myophore; the
wide anterior socket on RV has marginal canals (arrow) (b). Recumbent specimens
of Pachytraga paradoxa, i.e.,
those in which a coiled LV is involved in shell sediment support, are not very
common, and somewhat difficult to recognize in random sections; the best known
examples are from Portugal (Skelton & Masse,
1998). It is
worth noting that the specific position of the specimens from Portugal cannot be
ascertained. Moreover, in the specimen illustrated herein, in transverse cross
sections of the LV, the Dap is higher than the Ddv; this is the reverse in P.
paradoxa (Skelton & Masse,
1998). Therefore, the foregoing
questions the taxonomic significance of the recumbent morphology.
). Cross-section of LV is subtriangular with mid-ventral inner ridge
(a-b), RV (a) with anterior myophoral thickening (with small rounded canals).
The anterior tooth of LV projects into the anterior socket of RV (a); the RV has
a wide perimyophoral cavity and a broken (a) and a preserved (b) myophore; the
wide anterior socket on RV has marginal canals (arrow) (b). Recumbent specimens
of Pachytraga paradoxa, i.e.,
those in which a coiled LV is involved in shell sediment support, are not very
common, and somewhat difficult to recognize in random sections; the best known
examples are from Portugal (Skelton & Masse,
1998). It is
worth noting that the specific position of the specimens from Portugal cannot be
ascertained. Moreover, in the specimen illustrated herein, in transverse cross
sections of the LV, the Dap is higher than the Ddv; this is the reverse in P.
paradoxa (Skelton & Masse,
1998). Therefore, the foregoing
questions the taxonomic significance of the recumbent morphology.
|
|
Figure 20:
Pachytraga paradoxa (Pictet & Campiche). Combe des
pins. a-b) Paired longitudinal sections of a bivalve recumbent specimen, (a)
conjoined), (b) disjoined. See interpretation in the text. Scale bar 10 mm. |
Age. The rudist assemblage of Combe des Pins-La Madeleine identifies the Rustrel fauna of upper Barremian age (Martelites sarasini zone) but P. paradoxa also occurs in the overlying lower Aptian Saint-Montan Member.
Pachytraga aff. paradoxa (Pictet & Campiche, 1869)
(Fig. 21 ![]() )
)
Material. Four blocks (JPMA 18071-1, 2, 3 and 4) including each two sections from Saint-Montan.
Taxonomic position. The assignement to Pachytraga is based on the following characters: posterior myophore of RV flanked by a marginal perimyophoral cavity, LV with a wide posterior myophoral cavity, inner ventral ridge present in some sections.
The type
section of this form is represented by a RV (18071-2) with a single carina and
numerous rounded canals on posterior, anterior, and ventral sides (Fig.
21.a ![]() ).
The size is modest (average: Dap = 23 mm, Ddv = 39 mm but up to 50 mm) and the
shell displays an antero-posterior compression. There is a single, rounded
antero-ventral carina and a rounded ventral side. The anterior myophore of LV is
relatively thick, the perimyophoral cavity extends to the dorsal side, a strong
anterior tooth projects into the posterior myocardinal cavity of the opposite
valve (Fig. 21.a
).
The size is modest (average: Dap = 23 mm, Ddv = 39 mm but up to 50 mm) and the
shell displays an antero-posterior compression. There is a single, rounded
antero-ventral carina and a rounded ventral side. The anterior myophore of LV is
relatively thick, the perimyophoral cavity extends to the dorsal side, a strong
anterior tooth projects into the posterior myocardinal cavity of the opposite
valve (Fig. 21.a ![]() ). The inner ventral ridge is vigorous. Canals occur on the
dorsal zone but are poorly preserved. The antero-ventral sinus is widely open.
Sections poorly reflect the shape of the LV, one section (Fig.
21.b
). The inner ventral ridge is vigorous. Canals occur on the
dorsal zone but are poorly preserved. The antero-ventral sinus is widely open.
Sections poorly reflect the shape of the LV, one section (Fig.
21.b ![]() ) has an
arcuate morphology, whether this habit is specific or an expression of an
ecological morphotype is not clear. RV with a very large anterior socket,
ventral side convex bearing irregular rounded canals (Fig. 21.c
) has an
arcuate morphology, whether this habit is specific or an expression of an
ecological morphotype is not clear. RV with a very large anterior socket,
ventral side convex bearing irregular rounded canals (Fig. 21.c ![]() ), posterior
myophore thick, canals present in the perimyophoral cavity (Fig.
21.c
), posterior
myophore thick, canals present in the perimyophoral cavity (Fig.
21.c ![]() ), the
central tooth of moderate size bears adjacent canals. A juvenile and/or apical
section of RV shows the main cavities, among which the wide anterior socket (Fig.
21.d
), the
central tooth of moderate size bears adjacent canals. A juvenile and/or apical
section of RV shows the main cavities, among which the wide anterior socket (Fig.
21.d ![]() ), the juvenile lacks canals in the RV ventral side, as canals appear
progressively during ontogeny.
), the juvenile lacks canals in the RV ventral side, as canals appear
progressively during ontogeny.
Comparisons. Pachytraga aff. paradoxa differs from P. tubiconcha Astre, 1961, and P. carpathica Masse et al., 2023b, two species essentially lacking canals. Our specimens are close to Pachytraga paradoxa, and their average size is comparable to those of the larger specimens of the type species. The presence of a single rounded carina, a stronger LV anterior tooth, a wide RV anterior socket and the development of canals are the main differences with P. paradoxa, but P. gracilis is smaller and displays densely distributed minute canals (Masse et al., 2024b). The dissimilarities are mostly quantitative and many variable characters found in P. paradoxa tend to be more stable in the studied specimens, obviously closely related to the former species with which they coexist. We suggest that Pachytraga aff. paradoxa might be a new species characterized by its more pronounced canaliculation and some other morphological traits.
Age. Known only from its type locality P. aff. paradoxa is early Aptian.
|
|
Figure 21:
Pachytraga aff. paradoxa,
Saint-Montan (JPMA 18071). a) Longitudinal oblique section of LV showing the
myocardinal elements and the ventral inner ridge. b) Longitudinal section of a
LV showing the arcuate habit. c) Transverse section of a RV showing the
canaliculated architecture. d) Sections of RV focusing on canaliculation
(arrows). e) Transverse apical /juvenile section. Scale bar 10 mm. |
Genus Praecaprina Paquier, 1905
The myophoral organization of the genus defined by Paquier (1905) with Praecaprina varians as type species, matches the "caprinid type"of Mac Gillavry (see above). The genus is defined by the following: a row of rounded, subrectangular canals flanking the anterior perimyophoral and posterior myophoral cavity on LV, canals being absent on the ventral side and rare on the opposite valve. An inner ventral ridge is present in the LV, absent in the RV (Chartrousse, 1998). Some new species described herein will allow us to modify, to some extent, the foregoing definition (see below).
Praecaprina cf. varians Paquier, 1905
(Fig. 22 ![]() )
)
Material. We assign to this species two sections of bivalve specimens 302.002 and 302.017 from Cité du barrage, and an internal mold of a LV from Aigučze (JPMA 18066-1).
Specific attribution.
According to Chartrousse (1998), the LV of Praecaprina
varians and Praecaprina gaudryi
described by Paquier (1905) are much similar, but the organization
of the
RV is different. A nicely preserved and prepared LV of Praecaprina
varians from Saint-Montan illustrated by Paquier (1905: Pl. VIII,
fig. 7) is reproduced herein (Fig. 22.a ![]() ). The longitudinal sections of bivalves
from Cité du barrage show the canals on LV and the ventral inner ridge and RV
are without canals (Fig. 22.b-c
). The longitudinal sections of bivalves
from Cité du barrage show the canals on LV and the ventral inner ridge and RV
are without canals (Fig. 22.b-c ![]() ), and are assigned to P.
varians. The coiled internal mold of Aigučze
(Fig. 22.d
), and are assigned to P.
varians. The coiled internal mold of Aigučze
(Fig. 22.d ![]() ) shows
the tracks of the inner canaliculate architecture of a LV. The dimensions of our
specimens are slightly smaller than those of the type material: Dap = 60 mm, Ddv
= 40-50 mm (Paquier, 1905; Chartrousse,
1998).
) shows
the tracks of the inner canaliculate architecture of a LV. The dimensions of our
specimens are slightly smaller than those of the type material: Dap = 60 mm, Ddv
= 40-50 mm (Paquier, 1905; Chartrousse,
1998).
Age. Praecaprina varians has a significant record in the Rustrel fauna of the Monts-de-Vaucluse (Masse, 1976; Chartrousse, 1998). The specimens from Ardčche are early Aptian.
|
|
Figure 22:
Praecaprina cf. varians Paquier.
a) LV of Praecaprina varians from Saint-Montan, reproduced from Paquier, 1905
(Pl. VIII, fig. 7). b)
Longitudinal section of a bivalve specimen showing the coiling of the LV from
Cité du barrage (SU.PAL.2022.1.15.5.3). c) Longitudinal oblique sections of
conjoined valves from Cité du barrage (SU.PAL.2022.1.15.5.4). d) LV internal
mold showing the coiling and the tracks of the internal cavities (arrow), from
Aigučze (JPMA 18066-1). Scale bar 10 mm. |
Praecaprina gaudryi Paquier, 1905
(Fig. 23 ![]() )
)
Material.
Specimens are tranverse sections of LV showing the shell outline and internal organization: body cavity, myophoral cavity and canal pattern; and
there is a single section of RV displaying posterior canals. Sections of shells from
Trescouvieux (sample SU.PAL.2022.1.15.11.2) document an early dissolution of the
aragonite replaced by a pale beige, micritic infill (Fig. 23.a ![]() ). The sections
from Cité du barrage (SU.PAL.2022.1.15.5.5 and SU.PAL.2022.1.15.4.1) document
the usual diagenetic replacement of the aragonite by sparry calcite (Fig.
23.c-d
). The sections
from Cité du barrage (SU.PAL.2022.1.15.5.5 and SU.PAL.2022.1.15.4.1) document
the usual diagenetic replacement of the aragonite by sparry calcite (Fig.
23.c-d ![]() ).
).
Specific characters.
Dimensions are in the range of those of the type material: Dap = 40-60 mm, Ddv =
30-40 mm (Chartrousse, 1998). The LV of Praecaprina
gaudryi has an oval transverse outline with Ddv<Dap, the ventral side is
flat or slightly depressed, the anterior and posterior sides are convex and bear
subrectangular rounded canals, some are elongated radially. Posterior canals
tend to be larger and more numerous (up to 15) than the anterior (Chartrousse,
1998). A ventral inner ridge on LV is absent on the RV. The above characters are
close to those of P. varians.
Characters of the RV: the occurrence of several posterior canals typify P.
gaudryi (Fig. 23.d ![]() ). In addition as
shown by Chartrousse (1998), the walls flanking the canals are longer and
thinner in P. gaudryi than in P. varians.
). In addition as
shown by Chartrousse (1998), the walls flanking the canals are longer and
thinner in P. gaudryi than in P. varians.
Age. Praecaprina gaudryi is a relatively common species of the upper Barremian Rustrel fauna and has also been recorded in the lower Aptian from SE Spain (Chartrousse, 1998), its record in the Ardčche region documents its occurrence in the lower Aptian of SE France.
|
|
Figure 23:
Praecaprina gaudryi Paquier. a-c) LV sections showing the
posterior myophoral cavity and the canal pattern. a) section from
Trescouvieux,
canal walls with a micritic secondary structure. b) Transverse section near the
commissure (SU.PAL.2022.1.15.11.2). c) Transverse section pf a LV from Cité du
barrage (SU.PAL.2022.1.15.5.5). d) Posterior side of RV showing the
canaliculation (SU.PAL.2022.1.15.4.1). Scale bar 10 mm. |
Praecaprina paquieri n. sp.
(Fig. 24 ![]() )
)
Material. The two valves specimen (365) from Aigučze constitute the holotype (SU.PAL.2022.1.15.12.2 with serial sections 002/014).
Derivation of name. Dedicated to Victor-Lucien Paquier (1870-1911) for his contribution to our knowledge of rudist bivalves in the study region.
Generic assignment. Canals present on the LV posterior myophoral cavity and anterior perimyophoral cavity, ventral side lacking canals. Abundant canals on LV, rare on RV.
Diagnosis. Praecaprina with a subcircular transverse outline, having only 2-3 small RV anterior canals, coiled LV with thin subsquare canals on the anterior and posterior sides, thin LV anterior pediculate myophore and canaliculate posterior tooth.
Description. Subcylindrical
twisted RV, coiled LV (Fig. 24.a ![]() ). The dimensions Dap = 45-50 mm, Ddv average 45
mm; document an absence of dorso-ventral compression, the total length is 80 mm.
The RV has angular postero-ventral edges, the postero-ventral junction of the LV
is carinate (Fig. 24.b
). The dimensions Dap = 45-50 mm, Ddv average 45
mm; document an absence of dorso-ventral compression, the total length is 80 mm.
The RV has angular postero-ventral edges, the postero-ventral junction of the LV
is carinate (Fig. 24.b ![]() ). The number of LV perimyophoral anterior
canals is 7-8,
posterior canals 10-12. The posterior myophoral plate of RV is thin and
pediculate, somewhat protruding in the perimyophoral cavity (Fig.
24.c
). The number of LV perimyophoral anterior
canals is 7-8,
posterior canals 10-12. The posterior myophoral plate of RV is thin and
pediculate, somewhat protruding in the perimyophoral cavity (Fig.
24.c ![]() ). Two
anterior rounded canals are on the RV, near the commissure (Fig.
24.d
). Two
anterior rounded canals are on the RV, near the commissure (Fig.
24.d ![]() ).
The LV
canals are subsquared with rounded edges and thin walls, more or less festooned
outside (Fig. 24.e
).
The LV
canals are subsquared with rounded edges and thin walls, more or less festooned
outside (Fig. 24.e ![]() ). The preservation of the posterior row is bad, except for
the ventral large canals protruding ventrally (at the postero-ventral junction),
the anterior row is interrupted above the antero-ventral carina. The posterior
tooth bears fine tubes (Fig. 24.f
). The preservation of the posterior row is bad, except for
the ventral large canals protruding ventrally (at the postero-ventral junction),
the anterior row is interrupted above the antero-ventral carina. The posterior
tooth bears fine tubes (Fig. 24.f ![]() ).
).
Comparisons. The presence of LV anterior canals is a strong difference between P. tubifera n. sp. and P. vacua n. sp. The thin anterior LV myophore lacking tubular structures is also a typical feature. The overall canaliculation of P. paquieri n. sp. is close to that of the group of P. varians and P. gaudryi, but the lack of dorso-ventral compression and the small number of canals, especially on the RV, make the new species different. Actually the number of LV anterior canals (7-8) is comparable to that of P. gaudryi which has nevertheless a higher number (16-17) posterior ones as P. varians. The thin canal walls and their square-rounded shape somewhat festooned and the lack of any radial compression are different from that of other species, e.g., P. gaudryi and P. robusta n. sp. The thin pedunculate myophoral plate on RV is also a peculiar character. The taxonomic significance of the finely tubular posterior tooth (a character absent or not identified in all other species) is not clear because this trait may be due to a moderate or non-diagenetic alteration of the aragonitic tooth material, a common feature in well preserved still aragonitic, caprinid shells, e.g., Offneria murgensis (Masse, 1992).
Age. Praecaprina paquieri n. sp. belongs to the early Aptian assemblage of the Saint-Montan fauna.
|
|
Figure 24: Praecaprina paquieri n. sp. from
Aigučze, holotype
(SU.PAL.2022.1.15.12.2). a) View of the bivalve contour of the holotype.
b-f)
Transverse serial sections. b-d) Sections of RV (from the apex to the
commissure) showing the internal characters, notice in (c) the thin pediculated
posterior myophore, in (d) the canaliculated portion of the LV. e-f)
canaliculated (white arrows) sections of the LV (posterior side worn out)
showing the ventral inner ridge and the finely tubular posterior tooth (black
arrow). Scale bar 10 mm. |
Praecaprina tubifera n. sp.
(Fig. 25 ![]() )
)
Material. One section of LV (SU.PAL.2022.1.15.4.5) constitutes the holotype and is from Cité du barrage. The RV (SU.PAL.2022.1.15.4.6) from the same locality might also belong to the same species but the specific assignment is uncertain.
Generic assignment. Based on the LV posterior myophoral cavity flanked by canals, ventral side lacking canals (see discussion).
Derivation of name. From the tubular structure: Latin tubus (tube) and ferre (to bear) of the LV anterior shell margin.
Diagnosis. Praecaprina with posterior rectangular canaliculation adjacent to the myophoral cavity, and anterior fine, radial-tubular structure on LV.
Description. The LV section (Fig. 25.a ![]() ) is subrectangular,
dorso-ventrally compressed, with a rounded
posterior edge slightly protruding ventrally, the ventral side is depressed, has
an acute anterior carina and bears an inner radial ridge. Antero-posterior
dimensions range from 30 to 40 mm. Ten posterior rectangular canals with thick
radial walls thining towards the myophoral cavity, limited inward by inverted
festoons forming a thin boundary with the adjacent cavity. Myophoral cavity
large, the thin median wall connects ventrally with large rounded canals forming
an extension of the posterior row. The thick anterior side lacks canals but is
made of subradially oriented tubes. The cardinal area is poorly preserved.
) is subrectangular,
dorso-ventrally compressed, with a rounded
posterior edge slightly protruding ventrally, the ventral side is depressed, has
an acute anterior carina and bears an inner radial ridge. Antero-posterior
dimensions range from 30 to 40 mm. Ten posterior rectangular canals with thick
radial walls thining towards the myophoral cavity, limited inward by inverted
festoons forming a thin boundary with the adjacent cavity. Myophoral cavity
large, the thin median wall connects ventrally with large rounded canals forming
an extension of the posterior row. The thick anterior side lacks canals but is
made of subradially oriented tubes. The cardinal area is poorly preserved.
|
|
Figure 25:
Praecaprina tubifera n. sp. from Cité du barrage. a) Transverse
section of a LV. The white arrow points to the tubular structure
(SU.PAL.2022.1.15.4.5). b) Transverse section of a possible RV from the species
(SU.PAL.2022.1.15.4.6). Scale bar 10 mm. |
Comparisons. The fine radial structure of the LV anterior shell side of Praecaprina tubifera n. sp. is unique in the genus Praecaprina.
A section of RV (Fig. 25.b ![]() )
with anterior side having a porous tubular structure seemingly longitudinal,
extends to the dorsal side and surrounds the massive central tooth. The anterior
myophore is thick. On the posterior side the posterior myophore is thick, the
adjacent perimyophoral cavity bears 4-6 rounded canals, partly cut obliquely. The
body cavity is rounded, subquadrate and lacks any inner ridge. The organization of this valve mimics that of the LV but the shell is more robust, its
compression is antero-posterior instead of dorso-ventral and its cardinal area
very strong. Considering that this RV is the counterpart of the above LV is,
therefore, uncertain and the diagnosis of the species is consequently based on
the characters of the LV only.
)
with anterior side having a porous tubular structure seemingly longitudinal,
extends to the dorsal side and surrounds the massive central tooth. The anterior
myophore is thick. On the posterior side the posterior myophore is thick, the
adjacent perimyophoral cavity bears 4-6 rounded canals, partly cut obliquely. The
body cavity is rounded, subquadrate and lacks any inner ridge. The organization of this valve mimics that of the LV but the shell is more robust, its
compression is antero-posterior instead of dorso-ventral and its cardinal area
very strong. Considering that this RV is the counterpart of the above LV is,
therefore, uncertain and the diagnosis of the species is consequently based on
the characters of the LV only.
Age. Recorded with Praecaprina gaudryi, P. tubifera n. sp. is a member of the early Aptian Saint-Montan fauna.
Praecaprina vacua n. sp.
(Fig. 26 ![]() )
)
Material. Sections of LV from Cité du barrage, one with a portion of RV (SU.PAL.2022.1.15.5.6 is the holotype and SU.PAL.2022.1.15.5.7 are paratypes).
Derivation of name. From the open anterior perimyophoral cavity on LV.
Generic assignment. Based on the LV posterior myophoral cavity flanked by canals, ventral side lacking canals and anterior perimyophoral cavity.
Diagnosis. Praecaprina with LV anterior perimyophoral cavity open, non-canaliculated.
Description. The transverse
oblique section of LV holotype (Fig. 26.a ![]() ) shows the posterior myophoral cavity
flanked by radial rectangular canals with thin walls, the apparent elongation of
canals is due to the obliquity of the section (see also Fig. 26.b
) shows the posterior myophoral cavity
flanked by radial rectangular canals with thin walls, the apparent elongation of
canals is due to the obliquity of the section (see also Fig. 26.b ![]() ). The body
cavity is wide, has an inner ventral ridge, and the anterior perimyophoral
cavity lacks canals (Fig. 26.a-b, e
). The body
cavity is wide, has an inner ventral ridge, and the anterior perimyophoral
cavity lacks canals (Fig. 26.a-b, e ![]() ). On some sections
(Fig. 26.d
). On some sections
(Fig. 26.d ![]() ) posterior
canals of the LV look elliptical, subpyriform. The skeleton is relatively thin,
weakly calcified (e.g., Fig. 26.b
) posterior
canals of the LV look elliptical, subpyriform. The skeleton is relatively thin,
weakly calcified (e.g., Fig. 26.b ![]() ). Sections of
Figure 26.b
). Sections of
Figure 26.b ![]() (left) as
well as that of the holotype, suggest a coiled LV. The section of Figure
26.f
(left) as
well as that of the holotype, suggest a coiled LV. The section of Figure
26.f ![]() cuts a part of the RV which displays posterior canals, the section suggests that
this valve is more strongly calcified than the opposite. The available material
suggests relatively small dimensions, Dap being in the range of 20 mm and the
number of posterior canals on LV in the range of 12-13.
cuts a part of the RV which displays posterior canals, the section suggests that
this valve is more strongly calcified than the opposite. The available material
suggests relatively small dimensions, Dap being in the range of 20 mm and the
number of posterior canals on LV in the range of 12-13.
|
|
Figure 26: Praecaprina vacua n. sp. from Cité du barrage (SU.PAL.2022.1.15.5.6
and SU.PAL.2022.1.15.5.7). a) Oblique transverse section of the LV (holotype).
b) Section of a LV whorl. c) Tangential and longitudinal sections of
LV.
d)
Oblique transverse section of a LV, ventral part of the body cavity missing. e)
Transverse oblique section of a LV showing the inner ventral ridge. f) Section
cut in a bivalve specimen showing a canaliculated RV. The white arrow points to
the LV and RV canals. Scale bar 10 mm. |
Comparisons. The absence of canals on the LV anterior side of Praecaprina vacua n. sp. is unique in the genus Praecaprina.
Age. The species is recorded with Praecaprina gaudryi and Praecaprina tubifera n. sp., Praecaprina vacua n. sp., and belongs to the early Aptian Saint-Montan fauna.
Genus Caprina Orbigny
The genus is characterized by a row of numerous canals on LV, including the ventral side, the RV has a limited number of canals (Chartrousse, 1998).
Caprina douvillei Paquier, 1905
(Fig. 27 ![]() )
)
Material.
It consists of a single transverse, oblique section of LV from Trescouvieux
(362.002) (Fig. 27 ![]() ) but poorly preserved sections co-occur in this locality.
) but poorly preserved sections co-occur in this locality.
Description. The species was first described by Paquier (1905) and has been revised by Masse and Chartrousse (1997). The section shows the body and posterior myophoral cavities and the antero-ventral canals. The shell is relatively small, Dap = 10-25 mm, the LV is moderately salient and its beak modest, the ventral margin is convex. Canals are essentially present on the LV, they are mostly pyriform and they range in number from 35-40; they are present on the anterior, ventral and posterior sides of the LV but absent on RV.
Age. The species has a significant record in SE France where it was considered an early Aptian marker (Masse & Chartrousse, 1997) in beds corresponding actually with the Rustrel fauna, currently assigned to the upper Barremian. The age of its type level at Les Rimets (Vercors) remains uncertain (cf. Paquier, 1905), its presence in the Saint-Montan fauna testifies that C. douvillei extended into the early Aptian as formerly established in Spain by Skelton et al. (2010).
|
|
Figure 27: Caprina douvillei Paquier. Section of a LV showing the body
and posterior myophoral cavity, and the antero-ventral canaliculation (arrows).Trescouvieux. Scale bar 10 mm. |
Genus Offneria (Paquier, 1905) Masse, 1992
The genus was described by Paquier (1905) with Offneria rhodanica as type species, then emended by Masse (1992). It belongs to the group of Caprinidae having pallial canals on both valves. The RV canals possess a concave upward (towards the commissure) cupulate structure. The LV canals are radially oriented and transversally partitioned. In each valve they tend to be arranged in several rows in advanced species. The myophoral cavity of LV, teeth sockets and the body cavity are commonly tabulated (Chartrousse, 1998).
Offneria aff. rhodanica Paquier, 1905
(Fig. 28 ![]() )
)
Material. Two longitudinal sections of a RV (samples JPMA 18145a1 and a2) and a partly isolated LV obliquely cut (sample JPMA 18144).
Description. The RV is subconical, H = 130-150 mm, Ddv = 40-50 mm, Dap = 120 mm, hence with a
significant antero-posterior elongation, it displays an irregular growth mode
including abrupt corbelled swellings (Fig. 28.a-b ![]() ). The LV is curved and much
shorter H = 90 mm, and displays a ventral depression (Fig. 28.c-d
). The LV is curved and much
shorter H = 90 mm, and displays a ventral depression (Fig. 28.c-d ![]() ). Shell
thickness is ventrally in the range of 15 mm (LV) to 20 mm (RV). The shell is
therefore large, massive and robust. The longitudinal sections of RV, display
the canal files having transverse cupulate partitions with concavity mainly
apparent on wide canals (Fig. 28.e-f
). Shell
thickness is ventrally in the range of 15 mm (LV) to 20 mm (RV). The shell is
therefore large, massive and robust. The longitudinal sections of RV, display
the canal files having transverse cupulate partitions with concavity mainly
apparent on wide canals (Fig. 28.e-f ![]() ). On transverse sections the cupulate
canals look rounded (Fig. 28.g
). On transverse sections the cupulate
canals look rounded (Fig. 28.g ![]() ). The dual habit of the LV and RV canals,
especially the cupulate RV ones typify the genus Offneria
and its overall habit as well.
). The dual habit of the LV and RV canals,
especially the cupulate RV ones typify the genus Offneria
and its overall habit as well.
The dimensions are in the range of that of O. rhodanica even somewhat larger, the posterior radially arranged canals of LV are rather similar, but the dimensions of the RV are much higher and the growth mode different, i.e., similar to that of O. prebetica (Masse et al., 2015). The density of pyriform canals on LV is also higher than that of O. rhodanica. Nevertheless, differences with Offneria prebetica include the lack of any inner row of rounded canals on LV flanking the pyriform outer row, and a more pronounced ventral depression; but the morphology of O. aff. rhodanica is close to that of the Spanish form.
Age. Offneria rhodanica has a significant record in Provence (Masse, 1976) where it was considered an early Aptian marker (Masse & Chartrousse, 1997) in beds corresponding with the Rustrel fauna, currently assigned to the upper Barremian. The occurrence of O. aff. rhodanica in the Saint-Montan fauna testifies that Offneria, rather uncommon, extended into the early Aptian and represents a potential precursor of the late early Aptian O. prebetica from southern Spain.
|
|
Figure 28: Offneria aff. rhodanica Paquier
from Cité du barrage. Specimens JPMA 18144 and 18145. a) LV ventral side
showing the ventral depression, vd. b) LV dorsal side with broken apex,
commissure partlty preserved. c) transverse oblique section trough the
commissure showing the canaliculate network of the RV and the radial stucture of
the LV. d) close-up of the radial structure (partly crushed). Specimen JPMA
18145, RV. e-f) longitudinal sections showing the cylindro-conical habit with
corbelled swellings (arrow) and the cupulate canal structure. g) close-up of the
shell structure with cupules (cu). Scale bar 10 mm. |
Family Polyconitidae Mac Gillavry, 1937
Genus Horiopleura Douvillé, 1889
Horiopleura brevis Masse & Fenerci-Masse, 2017
(Fig. 29 ![]() )
)
The species differs from H. dumortieri (Matheron) by its morphology: D is higher than H, size, much larger, and on LV a stronger, more massive myophoral organization, moreover its stratigraphic distribution is younger (Masse & Fenerci-Masse, 2017).
Study material. An antero-posterior section from Aigučze (JPMA 18065-2), a part of LV from Cité du barrage (SU.PAL.2022.1.15.5.8) and a field photograph from Aigučze.
Description. The antero-posterior section (Fig. 29.a ![]() ) illustrates a
cylindro-conical RV with a
thickened outer calcitic shell layer, a low domal LV with a thin calcitic outer
shell layer. The RV posterior inner shell layer, formerly aragonitic, thickened
and concave upward in correspondence with the convex downward LV myophore. The
RV anterior myophore flat and sloping inward. The LV posterior myophore is
massive with a thick pedicle and a short perimyophoral cavity, the anterior
myophore is a bulge. The thick, massive shell conforms to that of the type
material of Monts de Vaucluse (Masse & Fenerci-Masse,
2017).
The field photograph (Fig. 29.b
) illustrates a
cylindro-conical RV with a
thickened outer calcitic shell layer, a low domal LV with a thin calcitic outer
shell layer. The RV posterior inner shell layer, formerly aragonitic, thickened
and concave upward in correspondence with the convex downward LV myophore. The
RV anterior myophore flat and sloping inward. The LV posterior myophore is
massive with a thick pedicle and a short perimyophoral cavity, the anterior
myophore is a bulge. The thick, massive shell conforms to that of the type
material of Monts de Vaucluse (Masse & Fenerci-Masse,
2017).
The field photograph (Fig. 29.b ![]() ) matches section 18065-2 but, is more proximal.
We interpret the close up of a LV posterior myophore (Fig. 29.c
) matches section 18065-2 but, is more proximal.
We interpret the close up of a LV posterior myophore (Fig. 29.c ![]() ) as an element of
the same species.
) as an element of
the same species.
Age. Horiopleura brevis a common member of the upper Barremian Rustrel fauna, is also common in the lower Aptian from Ardčche. It is worth noting that this species is known from the same stratigraphic interval in Southern Spain and extends to the upper Aptian in Central Europe and in the Pontides, which pertain to the Carpatho-Cimmerian Province (Masse & Fenerci-Masse, 2017; Masse et al., 2023a).
|
|
Figure 29: Horiopleura brevis Masse & Fenerci-Masse.
a)
Antero- posterior section of a bivalve specimen showing the myophoral organization, Aigučze
(JPMA 18065-2. b) Field photograph of a bivalve cross-cut
specimen showing the internal characters, Aiguéze. c) Close up of the
pediculate LV posterior myophore, Cité du barrage (SU.PAL.2022.1.15.5.8). Scale
bar 10 mm. |
Horiopleura helviorum Masse et al., 2026
(Fig. 30 ![]() )
)
The study material is from Aigučze.
The new species described recently (Masse et al., 2026), which bears the myophoral attributes of Horiopleura, is characterized by its small dimensions and a domal LV. It belongs to the goup of small species (Dap < 30 mm) having on both valves a tranverse flattening of the anterior myophores, similar to Debrunia.
Age. A rare species extending from the late Barremian to the early Aptian.
|
|
Figure 30:
Horiopleura helviorus Masse et
al. from Aigučze. a) Antero-posterior, longitudinal section of a
bivalve specimen, showing the polycontid organization of the myophores and the
convex LV (JPMA 18092). b) Ibidem, holotype, notice the posterior
cavity supporting the LV myophore (reproduced from Masse et al.,
2026). Scale bar 10 mm. |
Our study of rudist faunas from the upper Barremian and lower Aptian Urgonian limestones from Ardčche documents the presence of eighteen upper Barremian species and twenty five lower Aptian species. The upper Barremian, Rustrel fauna includes five new species, the lower Aptian Saint-Montan fauna includes seven new, exclusive species with a significant biostratigraphic potential.
New species described in the late Barremian include a new genus Valclusella, type species Valclusella valclusensis extending to the early Aptian, and two other new species Valclusella biconvexa a late Barremian-early Aptian form and Valclusella inflata restricted to the late Barremian. We describe Bicornucopina acuminata a new species tentatively ascribed to the genus Bicornucopina up to now unknown in western Europe, our new species extends to the early Aptian. Mathesia darderi is identified and discovered for the first time in SE France, from the upper Barremian to the lower Aptian.
The majority of early Aptian species (72%), are already present in the late Barremian. Requieniidae include five species inherited from the Rustrel or Orgon faunas, i.e., Matheronia virginiae, Matheronia aff. aptiensis, Requienia gryphoides, Requienia ammonia, and Toucasia carinata. In the Monopleuridae three species are inherited from the Rustrel or Orgon faunas: Monopleura varians, Debrunia occitanica, and Debrunia ucetiae. Members of the Caprinidae common with the Rustrel fauna are represented by: Pachytraga paradoxa, Praecaprina gr. gaudryi-varians, and Caprina douvillei. The Polyconitidae Horiopleura brevis and Horiopleura helviorum are also inherited from the Rustrel fauna.
The eight new, exclusively early Aptian forms include two Monopleuridae represented by Homopleura parva the smallest and youngest representative of the genus, and a new genus and species Myostyla ardescencis. Caprinidae record a set of new taxa, all belonging to genera already present in the Rustrel fauna, the new caprinid assemblage includes: Pachytraga aff. paradoxa, Praecaprina paquieri, Praecaprina tubifera, Praecaprina vacua and Offneria aff. rhodanica. Four to five more taxa pending description may be incorporated in this assemblage in the near future.
The fauna described earlier
from Saint-Montan by Paquier (1905) was represented by
Pachytraga paradoxa, Praecaprina
varians, P. gaudryi, Offneria
interrupta Paquier, and Offneria
rhodanica Paquier. Two forms, Pachytraga lapparenti and Offneria
intermedia, described by this author were subsequently considered as invalid
(Skelton & Masse, 1998; Chartrousse,
1998). This fauna,
sampled in the chalky limestones of the "Saint-Montant quarries" (Paquier,
1905) represents actually the Rustrel fauna. The chalky beds regarded herein as
equivalent to the U2 Member of the Monts-de-Vaucluse and assigned to the upper
Barremian, predate the Saint-Montan Member, which yields the lower Aptian fauna
of this study (Fig. 31 ![]() ).
).
From a palaeobiogeographical point of view the early Aptian fauna is typically west European and rooted in European ancestors, this is also true for the new genus Valclusella. Immigrants of the Mediterranean South Tethys margin are absent, the entry of Bicornucopina, the only evidence for a Carpatho-Cimmerian or Apulian influence, being an exception.
The foregoing shows that the Saint-Montan fauna includes eighteen inherited species from the antecedent late Barremian Orgon of Rustrel faunas, but about ten species of the Rustrel fauna are missing (see Masse et al., 2020). The missing taxa and the offshoot of seven new taxa, document an overwhelming novelty of the Saint-Montan fauna with regard to its predecessor. Drastic dissimilarities are also evident with their putative descendants of the Villaroya de los Pinares fauna from the Maestrat basin, having only a limited diversity, mostly inherited, and few (four to five taxa) novelties (Gili et al., 2016; Masse et al., 2020).
We consider the Saint-Montan rudist assemblage as a distinctive fauna belonging to the lower Aptian Urgonian-type faunal sequence of SE France and SE Spain formerly defined by Masse et al. (2020). This fauna fills a gap in between the Rustrel and Villaroya de los Pinares faunas, and is framed by two drowning boundary events dated from the latest Barremian Pseudocrioceras mazierei Zone below, and the mid lower Aptian Deshayesites forbesi p.p. Zone above (just below the Roloboceras hambrovi Subzone) (Pictet et al., 2015). The lower drowning is coeval with the Taxy black shales (Moullade et al., 2000; Machhour et al., 1998) a subevent (of the OAE1a event), which peaks in the lowermost part of the Pseudocrioceras mazieri Zone (Masse & Fenerci-Masse, 2011). This delimitation shows that the Oceanic anoxic Event OAE1a event (Selli event) known to predate the Villaroya de los Pinares fauna (Gili et al., 2016) postdates the Saint-Montan fauna (see above).
|
|
Figure 31:
Position of the lower Aptian Saint-Montan fauna into the Urgonian-type rudist
faunal sequence of SE France and Spain. |
The study late Barremian-early Aptian rudist faunas from Ardčche reveals some remarkable evolutionary aspects.
The oblique LV myophore of Myostyla n. gen., assumed to be anterior, is unique in the Monopleuridae; protruding downward lamellar myophores recorded in Debrunia Masse & Fenerci-Masse, 2009, are perpendicular to the commissural plane. The myophoral organization of our new genus also evokes a polyconitid arrangement, e.g., that of Polyconites Roulland (Skelton et al., 2010; Skelton, 2013a). Critical differences regard the orientation of the inner slope of the myophoral plate: outwards in the Polyconitidae and inwards in the case of Myostyla n. gen. Moreover in the Polyconitidae the LV myophoral plate is posterior and the anterior myophore, usually a crest, tends to be orthogonal to the commissure.
The protruding downward and inflation of the posterior myophore recorded in Valclusella n. gen. also evokes a polyconitid arrangement, e.g., that of Polyconites Roulland. This trend aligns with the development of the LV lamellar myophores recorded in advanced species of Debrunia, e.g., D. ucetiae and D. occitanica (Fenerci-Masse & Masse, 2010; Masse & Fenerci-Masse, 2018) present in the Rustrel and Saint-Montan faunas. The elongation of myophore was assumed to correlate with LV flattening (Fenerci-Masse & Masse, 2010) but this is not the case for Valclusella. We contend that the elongation of myophores in the Monopleuridae merely reflect parallel evolution rather than convergence of distinct but closely related lineages (see discussion in Masse & Fenerci-Masse, 2021).
The increasing canaliculation of Pachytraga aff. paradoxa relative to its potential ancestor Pachytraga paradoxa, takes place in the general trend through time for developing palleal cavities, i.e., myophoral or perimyophoral cavities, subsequently subdivided to form palleal canals. This trend has long been recognized in the Caprinidae (Mac Gillavry, 1937; Dechaseaux et al., 1969).
Praecaprina was first described by Paquier (1905) from our study region. Praecaprina andersoni and Praecaprina? pennyi defined by Harris and Hodson (1922) from the Barremian-Aptian of Trinidad, have been transferred to other genera. Praecaprina andersoni was assigned to the new genus Pseudocaprina first mentionned by Chartrousse (1998) then formally described by Chartrousse and Masse (2004), Praecaprina? pennyi being placed by Masse et al. (2013) in the genus Pantojaloria. Praecaprina was therefore considered absent from the New World (Chartrousse & Masse, 2004). The recognition of the genus Praecaprina and its species from Oman (Masse et al., 1998) is still imperfect and needs further studies, the same is true for representatives of this genus quoted by Masse (1985) from Apulia, on the southern margin of the Mediterranean Tethys.
The description of three new species of the genus associated with two preexisting ones testifies that the early Aptian p.p. coincides with a culmination in specific diversity of Praecaprina and that SE France appears as a privileged area in this respect.
Our new taxonomic discoveries urge us to propose an emendation of the definition of the genus Praecaprina. The absence of anterior canals on LV and the presence of a radial structure in Praecaprina tubifera n. sp. and the existence in Pr. vacua n. sp. of an open LV anterior perimyophoral cavity, depart from the classical organization of Praecaprina characterized by "rounded to oval pallial canals with simple radial plates in anterior, posterior and postero-ventral areas (Dechaseaux et al., 1969; Chartrousse, 1998).
The generic placement of the above species might be, therefore, questionable, but the overall organization: myophores, lack of ventral canals, inner ventral ridge, posterior canaliculation, fits a praecaprinid one. The foregoing suggests that the diagnosis of the genus Praecaprina has to be modified and must refer to the rectangular, piriform or rounded modes of canaliculation of the posterior perimyophoral cavity in LV, whereas on the anterior side canaliculation may be present or absent (as an open cavity), or represented by radial tubes.
We are aware of the provisional character of the foregoing points and we cannot prejudge about the overall evolutionary significance of the study of early Aptian rudist fauna before a more complete, quantitative and qualitative, overview of its composition will be achieved.
Upper Barremian-lower Aptian Urgonian-type platform carbonates from Ardčche yield diversified rudist faunas belonging to the families Requieniidae, Monopleuridae, Caprinidae, and Polyconitidae. The upper Barremian succession postdates the "Palorbitolina lenticularis guide level" (i.e., the Imerites giraudi zone) and is documented from two sites from the Ardeche river gorges. The associated rudists taxa with Pachytraga paradoxa, identify the Rustrel fauna, which includes the new genus Valclusella. Valclusella valclusensis n. sp. and Valclusella biconvexa n. sp. are common in the upper Barremian and the lower Aptian. The genus Bicornocupina, unknown so far from western Europe, is recorded and represented by a new species, B. acuminata with the same dual stratigraphic distribution. Mathesia darderi discovered for the first time in SE France, is a member of the Rustrel fauna.
The Saint-Montan limestones, the topmost member of the Urgonian Formation of the Ardčche region, are stratigraphically equivalent to the U3 member of the Monts-de-Vaucluse. Their age is early Aptian, and spans the Deshayesites oglanlensis and D. forbesi p.p. zones. The Saint-Montan limestones are bounded by two drowning discontinuities, the basal one dates from the latest Barremian Pseudocrioceras mazierei zone and the upper one of mid-early Aptian age is just antecedent to the Roloboceras hambrovi zone.
The early Aptian "Saint-Montan fauna" consists of 25 rudist species, of which 8 are new and 17 are inherited. Among them (65 %), were already present in the late Barremian. The inherited fauna consists of Requieniidae including five species from the Rustrel or Orgon faunas, i.e., Matheronia virginiae, Matheronia aff. aptiensis, Requienia gryphoides, Requienia ammonia, and Toucasia carinata. The Monopleuridae are represented by three species inherited from the Rustrel or Orgon faunas: Monopleura varians, Debrunia occitanica, and Debrunia ucetiae. Members of the Caprinidae common in the Rustrel fauna are represented by: Pachytraga paradoxa, Praecaprina gr. gaudryi-varians, and Caprina douvillei. The Polyconitidae Horiopleura brevis and Horiopleura helviorum, are also inherited from the Rustrel fauna. The new taxon is a single monospecific genus Myostyla, M. ardescencis. Seven exclusive new species belong to pre-existing genera include Homopleura parva, Pachytraga aff. paradoxa, Offneria aff. rhodanica, and three species of Praecaprina: P. tubifera, P. vacua and, P. paquieri. Variations in the canaliculation patterns of the left valve of Praecaprina suggest emending the diagnosis of the genus. It is worth noting that four or five new taxa, currently under study, will enhance the specificity of the assemblage, and may be incorporated in our inventory, in part provisory. The rudist assemblage has a west European character and possesses a significant biostratigraphic potential.
A remarkable evolutionary feature of the late Barremian-early Aptian taxa is the development of lamellar myophores in the Monopleuridae.
Among the inherited species from the antecedent late Barremian, up to ten species of the Rustrel fauna are missing. The foregoing and the offshoot of 8 new taxa, express an overwhelming novelty of the Saint-Montan fauna with regard to its predecessor. The Saint-Montan rudists represent a distinctive fauna belonging to the Barremian-lower Aptian Urgonian-type faunal sequence of SE France and SE Spain, and fills a gap between the Rustrel and the Villaroya de los Pinares faunas.
We are grateful to P.W. Skelton (formerly at Open University, Milton Keynes), R.W. Scott (Tulsa, Oklahoma), and an anonymous reviewer for their constructive comments, which have helped to improve the initial version of the manuscript. B. Granier is acknowledged for his important editorial contribution. We deeply thank Vincent Rommevaux (Centre de Recherche en Paléontologie- Paris, Muséum National d'Histoire Naturelle) for the important work of cutting and polishing most of the rudist sections described here.
Astre G. (1933).- Sur les petits Agria tubuleux de l'Urgo-Aptien.- Bulletin de la Société Géologique de France (5e Série), Paris, t. III, p. 99-105.
Astre G. (1961).- Pachytraga tubuleux du Barrémien du Doubs.- Bulletin de la Société d'Histoire Naturelle de Toulouse, t. 96, fasc. 3-4, p. 205-222.
Bastide F. (2014).- Synthčse de l'évolution de la plateforme urgonienne (Barrémien tardif ŕ Aptien précoce) du Sud-Est de la France : Facičs, micropaléontologie, géochimie, géométries, paléotectonique et géomodélisation.- Thčse, Sciences de la Terre, Université de Grenoble ; Université de Lausanne, 417 p.
Bilotte M., Bousquet J.-P. & Debroas E.-J. (2017).- Gisements pyrénéens d'Offneria rhodanica Paquier, 1905 et de Caprina choffati Douvillé, 1898 (Rudistes).- Bulletin de la Société d'Histoire Naturelle de Toulouse, t. 153, p. 67-75.
Blanc J.-J. & Masse J.-P. (1965).- L'Urgonien du chaînon de La Fare-Lançon (B.-du-Rhône).- Comptes Rendus sommaires de la Société géologique de France (7e Série), Paris, fasc. 9, p. 292-293.
Chartrousse A. (1998).- Les Caprinidae (Rudistes) du Crétacé inférieur.- Thčse de Doctorat, Université de Provence (Aix-Marseille I), Paléontologie, 281 p.
Chartrousse A. & Masse J.-P. (2004).- Revision of the early Aptian Caprinidae (Rudist bivalves) of the New World. Evolutionary and palaeobiogeographic implications.- Courier Forschungsinstitut Senckenberg, Band 247, p. 19-34.
Clavel B., Charollais J., Busnardo R., Granier B., Conrad M., Desjacques P. & Metzger J. (2014).- La plate-forme carbonatée urgonienne (Hauterivien supérieur-Aptien inférieur) dans le Sud-Est de la France et en Suisse : Synthčse.- Archives des Sciences, Genčve, Band 67, Heft 1, p. 1-97. URL: https://www.e-periodica.ch/cntmng?pid=ads-004:2014:67::192
Clavel B., Conrad M.A., Busnardo R., Charollais J. & Granier B. (2013).- Mapping the rise and demise of Urgonian platforms (Late Hauterivian-Early Aptian) in southeastern France and the Swiss Jura.- Cretaceous Research, vol. 39, p. 29-46.
Cotillon P., Ferry S., Busnardo R., Lafarge D. & Renaud B. (1979).- Synthčse stratigraphique et paléogéographique sur les facičs urgoniens du Sud de l'Ardčche et du Nord du Gard (France).- Géobios, Villeurbanne, Mémoire spécial 3, p. 121-139.
Császár G., Szinger B. & Piros O. (2013).- From continental platform towards rifting of the Tisza Unit in the late Triassic to early Cretaceous.- Geologica Carpathica, Bratislava, vol. 64 no. 4, p. 279-290. URL: https://geologicacarpathica.com/browse/archive/?journal_article_no=8639
Császár G. & Turnsek D. (1996).- Vestiges of atoll-like formations in the Lower Cretaceous of the Mecsek Mountains, Hungary.- Cretaceous Research, vol. 17, no. 4, p. 419-442.
Czabalay L. (1968).- Mecseki kréta Pachyodonták (Pachyodontes crétacés de la Montagne Mecsek).- Magyar Allami Földtani Intézet évi Jelentése, Budapest, 1968. Évről, p. 185-211. URL: https://epa.oszk.hu/02900/02934/00120/pdf/EPA02934_mafi_evi_jel_1968_185-211.pdf
Dechaseaux C., Cox L.R., Coogan A.H. & Perkins B.F. (1969).- Superfamily Hippuritacea Gray, 1848. In: Moore R.C. (ed.), Mollusca 6, Bivalvia 2.- Treatise on Invertebrate Paleontology, Boulder - CO, Lawrence - KS, Part N, N749-N817.
Delanoy G., Baudouin C., Pictet A., Moreno-Bedmar J., Frau C. & Matrion B. (2022).- The genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949 (Ammonoidea, Ancyloceratina, Douvilleiceratidae), from the Lower Aptian of Ardčche (SE France) - Taxonomic and biostratigraphic implications.- Carnets Geol., Madrid, vol. 22, no. 2, p. 7-109. DOI: 10.2110/carnets.2022.2202
Douvillé H. (1887).- Sur quelques formes peu connues de la Famille des Chamidés.- Bulletin de la Société géologique de France (3e Série), Paris, t. XV, p. 756-802.
Douvillé H. (1915).- Les Réquiénidés et leur évolution.- Bulletin de la Société Géologique de France (4e Série), Paris, t. XIVe, p. 383-389.
Douvillé H. (1918).- Le Barrémien supérieur de Brouzet. IIIe partie : Les rudistes.- Mémoires de la Société Géologique de France (Paléontologie), Paris, no. 52, t. XXII, p. 5-19 (IV Pls.). URL: https://patrimoine.sorbonne-universite.fr/idurl/1/2889
Fenerci-Masse M. & Masse J.-P. (2010).- Debrunia occitanica nov. sp. (Monopleuridae) from the early Aptian of SE France.- Turkish Journal of Earth Sciences, Ankara, vol. 19, no. 5, p. 573-581.
Fenerci-Masse M., Masse J.-P., Kołodziej B., Ivanov M. & Idakieva V. (2011).- Mathesia darderi (Astre) (Bivalvia, Hippuritoidea, Monopleuridae): Morphological, biogeographical and ecological changes in the Mediterranean domain during the late Barremian-Albian.- Cretaceous Research, vol. 32, no. 4, p. 407-421.
Frau C. & Delanoy G. (2022).- Taxonomic notes on some Barremian-Aptian cephalopods from the Station de Cassis section and surrounding sites, Bouches-du-Rhône, southern France.- Strata, Gaillac, série 2, vol. 58, p. 1-45.
Frau C., Tendil A.J.B., Lanteaume C., Masse J.-P., Pictet A., Bulot L.G., Luber T.L., Redfern J., Borgomano J.R., Léonide P., Fournier F. & Massonnat G. (2018).- Late Barremian-early Aptian ammonite bioevents from the Urgonian-type series of Provence, southeast France: regional stratigraphic correlations and implications for dating the peri-Vocontian carbonate platforms.- Cretaceous Research, vol. 90, p. 222-253.
Gili E., Skelton P.W., Bover-Arnal T., Salas R., Obrador A. & Fenerci-Masse M. (2016).- Depositional biofacies model for post-OAE1a Aptian carbonate platforms of the western Maestrat Basin (Iberian Chain, Spain).- Palćogeography, Palćoclimatology, Palćoecology, vol. 453, p. 101-114.
Goldfuss A. (1837).- Petrefacta Germanić tam ea, quae in Museo Universitatis Regiae Borussicae Fridericiae Wilhelmiae Rhenanae servantur quam alia quaecunque in Museis Hoeninghusiano, Muensteriano aliisque extant, iconibus et descriptionibus illustrata.- Zweiter Teil, Arnz & Cie., Düsseldorf, p. 141-224 (Pls. CXXII-CXLVI).
Granier B., Clavel B., Busnardo R., Charollais J., Desjacques P. & Bert D. (2021).- Biostratigraphic distribution of orbitolinids in the ammonite biozones (Urgonian platform of southeastern France). Part 2: Barremian p.p.- Carnets Geol., Madrid, vol. 21, no. 18, p. 399-521. DOI: 10.2110/carnets.2021.2118
Gras A. (1854).- Catalogue des corps organisés fossiles qui se rencontrent dans le département de l'Isčre.- Bulletin de la Société Statistique, des Sciences naturelles et des Arts Industriels du département de l'Isčre (2e Série), Grenoble, t. IIe, p. 1-54. URL: https://gallica.bnf.fr/ark:/12148/bpt6k5810111b/f7.item
Harris G.D. & Hodson F. (1922).- The rudists of Trinidad.- Palaeontographica Americana, Ithaca - NY, no. 14, p. 119-162.
Hofmann K. & Vadász E. (1912).- A Mecsekhegység középső-neokom rétegeinek kagylói.- Magyar Állami Földtani Intézet évkönyve, 20. köt., 5. füz., p. 189-226. URL: https://epa.oszk.hu/03200/03274/00178/pdf/
Kutassy A. (1934).- Pachyodonta Mesozoica. In: Quenstedt W. (ed.), Fossilium Catalogus. I. Animalia.- W. Junk, Gravenhage, pars 68, 202 p.
Lafarge D. (1978).- Étude géologique du plateau de Saint-Remčze, Ardčche : stratigraphie, cartographie, sédimentologie, tectonique.- Thčse de 3e cycle, Université Claude Bernard, Lyon, 119 p.
Leenhardt F. (1883).- Étude géologique de la région du Mont-Ventoux.- Thčse de doctorat, Université de Montpellier, 273 p.
Léonide P., Borgomano J., Masse J.-P. & Doublet S. (2012).- Relation between stratigraphic architecture and multi-scale heterogeneities in carbonate platforms: The Barremian-Lower Aptian of the Monts de Vaucluse, SE France.- Sedimentary Geology, vol. 265-266, p. 87-109.
Löser H. & Ferry S. (2006).- Coraux du Barrémien du Sud de la France (Ardčche et Drôme).- Géobios, Villeurbanne, vol. 39, no. 4, p. 469-489.
Lory C. (1846).- Études sur les terrains secondaires des Alpes, dans les environs de Grenoble.- Thčse de doctorat, Université de Paris, 135 p. URL: https://patrimoine.sorbonne-universite.fr/idurl/1/2295
Mac Gillavry H.J. (1937).- Geology of the province of Camaguey, Cuba with revisional studies of rudist paleontology.- Thčse de doctorat, Université d'Utrecht, 168 p. URL: https://dspace.library.uu.nl/handle/1874/306891
Machhour L., Masse J.-P., Oudin J.-L., Lambert B. & Lapointe P. (1998).- Petroleum potential of dysaerobic carbonate source rocks in an intrashelf basin: The Lower Cretaceous of Provence, France.- Petroleum Geosciences, vol. 4, no. 2, p. 139-146.
Mainelli M. (1996).- The rudist Mathesia tertiicolloquiirudistarum n. gen. n. sp. in the upper Aptian of Serra Sbregavitelli, Matese, south Apennines, Italy.- Revista Mexicana de Ciencias Geológicas, Querétaro, vol. 12, no. 2, p. 201-210. URL: https://rmcg.geociencias.unam.mx/index.php/rmcg/article/view/1193
Martínez-Rodríguez R., Castro J.-M., Gea G.A. de, Nieto L.M., Ruiz-Ortiz P. & Skelton P.W. (2025).- Early Aptian development and OAE 1a-linked demise of a carbonate platform in the Western Tethys: Lower Cretaceous of Sierra Mariola (South Iberian Paleomargin, SE Spain).- Cretaceous Research, volume 167, article 106032, 25 p.
Masse J.-P. (1976).- Les calcaires urgoniens de Provence (Valanginien-Aptien inférieur). Stratigraphie, paléontologie, les paléoenvironnements et leur évolution.- Thčse Université Aix-Marseille II, 445 p.
Masse J.-P. (1985).- Paléobiogéographie des rudistes du domaine péri-méditerranéen ŕ l'Aptien inférieur.- Bulletin de la Société Géologique de France (8e Série), Paris, t. I, no. 5, p. 715-721.
Masse J.-P. (1992).- Les rudistes de l'Aptien inférieur d'Italie continentale : Aspects systématiques, stratigraphiques et paléobiogéographiques.- Geologica Romana, vol. XXVIII, p. 243-260.
Masse J.-P. (1993).- Valanginian to early Aptian carbonate platforms from Provence (SE France). In: Simo A., Scott R.W. & Masse J.-P. (eds.), Cretaceous carbonate platforms.- American Association of Petroleum Geologists, Boulder - CO, Memoir 56, p. 363-374.
Masse J.-P. (1996).- Lower Cretaceous rudist biostratigraphy of southern France: A reference for Mesogean correlations.- Revista Mexicana de Ciencias Geológicas, Querétaro, vol. 12, no. 2, p. 236-256. URL: https://rmcg.geociencias.unam.mx/index.php/rmcg/article/view/1196
Masse J.-P. (2002).- Modifications morphologiques et anatomiques chez les Requieniidae (Rudistes) du Crétacé inférieur. In: Sladić-Trifunović M. (ed.), Proceedings of the 1st International Conference on Rudists (Beograd, 1988).- Serbian Geological Society Special Publication, 2, Beograd (1991), p. 155-171.
Masse J.-P. (2003).- Integrated stratigraphy of the Lower Aptian and applications to carbonate platforms: A state of the art. In: Gili E., Negra M. & Skelton P.W. (eds), North African Cretaceous carbonate platform systems.- Kluwer Academic Publishers, Bonn, p. 203-214.
Masse J.-P., Arias C. & Vilas L. (1998).- Lower Cretaceous rudist faunas of southeast Spain: An overview. In: Masse J.-P. & Skelton P.W. (eds.), Quatričme Congrčs International sur les Rudistes.- Géobios, Villeurbanne, Mémoire Spécial 22, p. 193-210.
Masse J.-P. & Chartrousse A. (1997).- Les Caprina (Rudistes) de l'Aptien inférieur d'Europe occidentale : Systématique, biostratigraphie et paléobiogéographie.- Géobios, Villeurbanne, vol. 30, no. 6, p. 797-809.
Masse J.-P., Chartrousse A. & Borgomano J. (1998).- The Lower Cretaceous (Upper Barremian-Lower Aptian) caprinid rudists from Oman. In: Masse J.-P. & Skelton P.W. (eds.), Quatričme Congrčs International sur les Rudistes.- Géobios, Villeurbanne, Mémoire Spécial 22, p. 211-233.
Masse J.-P. & Fenerci-Masse M. (2009).- Debrunia, a new Barremian genus of petalodontid Monopleuridae (Bivalvia, Hippuritoidea) from the Mediterranean region.- Palaeontology, London, vol. 52, no. 6, p. 1363-1372. DOI: 10.1111/j.1475-4983.2009.00912.x
Masse J.-P. & Fenerci-Masse M. (2010).- Mathesia Mainelli (Hippuritacea, Monopleuridae) from the late Aptian-Albian of the Mediterranean region, a revision.- Turkish Journal of Earth Sciences, Ankara, vol. 19, no. 5, p. 543-556. DOI: 10.3906/yer-0901-11
Masse J.-P. & Fenerci-Masse M. (2011).- Drowning discontinuities and stratigraphic correlation in platform carbonates. The late Barremian-early Aptian record of southeast France.- Cretaceous Research, vol. 32, no. 6, p. 659-684.
Masse J.-P. & Fenerci-Masse M. (2013a).- Stratigraphic updating and correlation of Late Barremian-Early Aptian Urgonian successions and their marly cover, in their type region (Orgon-Apt, SE France).- Cretaceous Research, vol. 39, p. 17-28.
Masse J.-P. & Fenerci-Masse M. (2013b).- Bioevents and palaeoenvironmental changes in carbonate platforms: The record of Barremian "Urgonian" limestones of SE France.- Palćogeography, Palćoecology, Palćoclimatology, vol. 386, p. 637-651.
Masse J.-P. & Fenerci-Masse M. (2013c).- Drowning events, development and demise of carbonate platforms and controlling factors: the Late Barremian-Early Aptian record of southeast France.- Sedimentary Geology, vol. 298, p. 28-52.
Masse J.-P. & Fenerci-Masse M. (2017).- Taxonomy and stratigraphy of late Barremian-Albian species of Horiopleura Douvillé (Hippuritida, Polyconitidae).- Cretaceous Research, vol. 76, p. 53-80.
Masse J.-P. & Fenerci-Masse M. (2018).- Taxonomy and stratigraphy of Lower Cretaceous species of Debrunia Masse and Fenerci-Masse (Hippuritida, Monopleuridae) of the Mediterranean region.- Cretaceous Research, vol. 84, p. 32-61.
Masse J.-P. & Fenerci-Masse M. (2019).- Homopleura new genus (Hippuritida, Monopleuridae) and the origin of the Polyconitidae.- Cretaceous Research, vol. 97, p. 107-124.
Masse J.-P. & Fenerci-Masse M. (2020).- Taxonomy and biostratigraphy of early Cretaceous species of the genus Monopleura Matheron (Hippuritida, Monopleuridae) of the Mediterranean region.- Cretaceous Research, vol. 108, article 104287, 28 p.
Masse J.-P. & Fenerci-Masse M. (2021).- Convergent and parallel evolutionary traits in Early Cretaceous rudist bivalves (Hippuritidina).- International Journal of Paleobiology and Paleontology, Novi - MI, vol. 4, no. 1, article 000120, 8 p. URL: https://medwinpublishers.com/IJPBP/convergent-and-parallel-evolutionary-traits-in-early-cretaceous-rudist-bivalves-hippuritidina.pdf
Masse J.-P., Fenerci-Masse M., Arias C. & Vilas L. (2015).- Description of a new species of Offneria (Hippuritida, Caprinidae) from the lower Aptian of southeast Spain. Stratigraphic, evolutionary, palaeobiogeographic and palaeoenvironmental implications.- Cretaceous Research, vol. 53, p. 153-166.
Masse J.-P., Fenerci-Masse M. & Bucur I.I. (2023a).- The Lower Cretaceous Carpatho-Cimmerian bioprovince: The contribution of rudist bivalves (Hippuritida).- Cretaceous Research, vol. 144, article 105448, 15 p.
Masse J.-P., Fenerci-Masse M. & Bucur I.I. (2023b).- Pachytraga carpathica nov. sp. (Hippuritida, Caprinidae) from the Carpatho-Balkanic region (Romania). Evolutionary and biogeographic implications for the genus Pachytraga Paquier.- Cretaceous Research, vol. 149, article 105550, 8 p.
Masse J.-P., Fenerci-Masse M. & Bucur I.I. (2024a).- New species of the Lower Cretaceous genus Matheronia Munier-Chalmas (Bivalve Hippuritida) in Romania.- Acta Palaeontologica Romaniae, Bucharest, vol. 20, no. 1, p. 3-15.
Masse J.-P., Fenerci-Masse M. & Mitchell S. (2013).- Revision of Praecaprina? pennyi (Harris and Hodson) (Caribbean caprinid rudist): Its placement in Pantojaloria Alencaster and Pantoja-Alor; taxonomic, stratigraphic and paleobiogeographic implications.- Caribbean Journal of Earth Sciences, Kingston - JM, vol. 45, p. 99-105. URL: http://caribjes.com/CJESpdf/CJES45-10-MassePraecaprina.pdf
Masse J.-P., Fenerci-Masse M., Platel J.-P., Frau C. & Léonide P. (2026).- New species of the genus Horiopleura (Bivalvia, Hippuritida, Polyconitidae) from the Barremian-lower Aptian deposits of the Mediterranean region: Evolutionary implications.- Acta Palaeontologica Romaniae, Bucharest, vol. 21, no. 2 (2025), p. 41-52.
Masse J.-P., Frau C. & Fenerci-Masse M. (2024b).- Pachytraga gracilis sp. nov. (Hippuritida, Caprinidae) from the Barremian of the Sub-Alpine region (SE France). Evolutionary and biogeographic implications for the genus Pachytraga Paquier.- Cretaceous Research, vol. 153, article 105706, 10 p.
Masse J.-P., Frau C., Tendil A.J.B. & Fenerci-Masse M. (2020).- Evidence for three successive upper Barremian-lower Aptian rudist faunas in the Urgonian-type deposits of South-Eastern France and their stratigraphic value.- Cretaceous Research, vol. 115, article 104561, 15 p.
Masse J.-P., Hadach F., Fenerci-Masse M., Algouti Ahm. & Algouti Abd. (2019).- The late Aptian requieniid rudist Pseudotoucasia catalaunica Astre from the Mediterranean region with emphasis on Moroccan occurences of the species.- Journal of African Earth Sciences, vol. 151, p. 548-554.
Matheron P. (1842).- Catalogue méthodique et descriptif des corps organisés fossiles du département des Bouches-du-Rhône et lieux circonvoisins.- Carnaud et Fils, Marseille, 269 p.
Matheron P. (1878).- Recherches paléontologiques dans le Midi de la France, ou étude sur les animaux fossiles découverts dans cette région présentant la description et la figure des espčces nouvelles, douteuses ou peu connues avec l'énumération méthodique des corps organisés fossiles qui le accompagnent dans leurs gisements stratigraphiques.- Livraisons 1 & 2, chez l'auteur, Marseille, iv + 7 p. (41 Pls.).
Moullade M., Masse J.-P., Tronchetti G., Kuhnt W., Ropolo P., Bergen J.A., Masure E. & Renard M. (2000).- Le stratotype historique de l'Aptien inférieur (région de Cassis-La Bédoule, SE France) : Synthčse stratigraphique.- Géologie Méditerranéenne, Marseille, t. XXV, no. 3/4, p. 289-298. URL: https://www.persee.fr/doc/geolm_0397-2844_1998_num_25_3_1641
Munier-Chalmas H. (1873).- Prodrôme d'une classification des rudistes.- Journal de Conchyologie, Paris, vol. 13, no. 3, p. 71-75.
Paquier V. (1896).- Sur quelques rudistes nouveaux de l'Urgonien.- Travaux du laboratoire de Géologie, Université de Grenoble, vol. 4, no. 1, p. 73-77.
Paquier V. (1900).- Recherches géologiques dans le Diois et le Baronnies orientales.- Allier frčres, Grenoble, 395 p.
Paquier V. (1903).- Les rudistes urgoniens. Premičre partie.- Mémoire de la Société géologique de France (Paléontologie), Paris, no. 29, t. XII, 46 p. URL: https://patrimoine.sorbonne-universite.fr/idurl/1/2821
Paquier V. (1905).- Les rudistes urgoniens. Deuxičme partie. Série inverse.- Mémoire de la Société géologique de France (Paléontologie), Paris, no. 29, t. XIII, p. 47-102.
Pictet A. & Delanoy G. (2017).- The Chabert formation: A newly defined stratigraphic unit of late early Aptian age in the southern Ardčche, SE France.- Archives des Sciences, Genčve, Band 69, Heft 1, p. 3-28. URL: https://www.e-periodica.ch/digbib/view?pid=ads-004%3A2017%3A69%3A%3A8
Pictet A., Delanoy G., Adatte T., Spangenberg J.E., Baudouin C., Boselli P., Boselli M., Kindler P. & Föllmi K.B. (2015).- Three successive phases of platform demise during the early Aptian and their association with the oceanic anoxic Selli episode (Ardčche, France).- Palćogeography, Palćoecology, Palćoclimatology, vol. 418, p. 101-125.
Pictet F.J. & Campiche G. (1869).- Description des fossiles du terrain Crétacé des environs de Sainte-Croix.- Matériaux pour la Paléontologie Suisse, Genčve, 4e partie, 2e livraison, 352 p. (Pls. CXL-CXCIV). URL: https://books.google.fr/books?id=xgJN_2Twl9YC&hl=fr&pg=PA48#v=onepage&q&f=false
Quesne D. (1996, unpublished).- Corrélations de détail entre les calcaires urgoniens et les facičs marno-calcaires du bassin subalpin (Barrémien, France sud-est).- Thčse de doctorat, Université Claude Bernard Lyon-I, 2 volumes, iii + 99 p.
Reboulet S., Jaap K. (reporters), Barragán R., Company M., González-Arreola C., Lukeneder A., Raisossadat S. N., Sandoval J., Szives O., Tavera J. M., Vašícek Z. & Vermeulen J. (2009).- Report on the 3rd International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the "Kilian Group" (Vienna, Austria, 15th April 2008).- Cretaceous Research, vol. 30, no. 2, p. 496-502.
Reboulet S., Szives O., Aguirre-Urreta B., Barragan R., Company M., Idakieva V., Ivanov M., Kakabadze M.V., Moreno-Bedmar J.A., Sandoval J., Baraboshkin E.J., Caglar M.K., Fözy I., Gonzales-Arreola C., Kenjo S., Lukeneder A., Raisossadat S.N., Rawson P.F. & Tavera J.M. (2014).- Report on the 5th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Ankara, Turkey, 31st August, 2013).- Cretaceous Research, vol. 50, p. 126-137.
Rineau V., Masse J.-P. & Villier L. (2020).- A new cladistic insight on comparative anatomy and phylogeny of rudists (Bivalvia, Hippuritida).- Journal of Systematic Palaeontology, vol. 18 no. 15, p. 1243-1297.
Scott R.W., Wan X., Sha J. & Wen S.-X. (2010).- Rudists of Tibet and the Tarim Basin, China: significance to Requieniidae phylogeny.- Journal of Palaeontology, Cambridge (UK), vol. 84, no. 3, p. 444-465.
Skelton P.W. (1976, unpublished).- Investigations into the palaeobiology of rudists.- PhD Thesis, Oxford University, 263 p.
Skelton P.W. (2004).- Oedomyophorus shaybahensis, a new genus and species of caprinid (?) rudist from the lower aptian Shu'aiba formation of Eastern Saudi Arabia.- Courier Forschungsinstitut Senckenberg, Band 247, p. 35-47.
Skelton P.W. (2013a).- Rudist classification for the revised Bivalvia volumes of the 'Treatise on Invertebrate Paleontology'.- Caribbean Journal of Earth Sciences, Kingston - JM, vol. 45, p. 9-33. URL: https://caribjes.com/CJESpdf/CJES45-04-SkeltonClassification.pdf
Skelton P.W. (2013b).- Rudist classification: nomenclatural correction of "Suborder Radiolitidina Skelton, 2013" to "Suborder Hippuritidina Newell, 1965".- Caribbean Journal of Earth Sciences, Kingston - JM, vol. 45, p. 34. URL: https://caribjes.com/CJESpdf/CJES45-04a-SkeltonClassification.pdf
Skelton P.W., Castro J.M. & Ruiz-Ortiz P.A. (2019).- Aptian carbonate platform development in the Southern Iberian palaeomargin (Prebetic of Alicante, SE Spain).- BSGF, Earth Sciences Bulletin, Paris, vol. 190, article 3, 19 p. DOI: 10.1051/bsgf/2019001
Skelton P.W., Gili E., Bover-Arnal T., Salas R. & Moreno-Bedmar J.A. (2010).- A new species of Polyconites from the Lower Aptian of Iberia and the early evolution of polyconitid rudists.- Turkish Journal of Earth Sciences, Ankara, vol. 19, no. 5, p. 557-572. URL: https://journals.tubitak.gov.tr/earth/vol19/iss5/3/
Skelton P.W. & Masse J.-P. (1998).- Revision of the Lower Cretaceous rudist genera Pachytraga Paquier and Retha Cox (Bivalvia, Hippuritacea) and the origins of the Caprinidae. In: Masse J.-P. & Skelton P.W. (eds.), Quatričme Congrčs International sur les Rudistes.- Géobios, Villeurbanne, Mémoire Spécial 22, vol. 31, suppl. 1, p. 331-370.
Stein M., Arnaud-Vanneau A., Adatte T., Fleitmann D., Spangenberg J.E. & Föllmi K.B. (2012).- Palaeoenvironmental and palaeoecological change on the north Tethyan carbonate platform during the late Barremian to earliest Aptian.- Sedimentology, vol. 59, p. 939-963.
Tendil A.J.B., Frau C., Léonide P., Fournier F., Borgomano J.R., Lanteaume C., Masse J.-P., Massonnat G. & Rolando J.-P. (2018).- Platform-to-basin transition of the Urgonian carbonate platform in Provence (Barremian-Aptian, SE France): New insights into the regional to global factors controlling the local stratigraphic architecture.- Cretaceous Research, vol. 91, p. 382-411.