◄ Carnets Geol. 26 (2) ►
![]()
Outline:
[1. Introduction][2. Geographical and geological setting]
[3. Material and method]
[4. Palynological results] [5. Discussion]
[6. Conclusions]
[Bibliographic references] and ...[Plates]
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
naima.berry-etu@etu.univh2c.ma
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
nadia.barhoun@univh2c.ma
Sorbonne Université, CNRS-INSU, Institut des Sciences de la Terre Paris,
ISTEP, Laboratoire Evolution et Modélisation des Bassins Sédimentaires UMR 7193, 75005 Paris (France)
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
Cadi Ayyad University, Faculty of Sciences Semlalia, Geology Department, Laboratory of Water Sciences, Microbial Biotechnology, and Sustainability of Natural Resources, Bd. Prince My
Abdellah, P.O. Box 2390, 40000 Marrakech (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben
M'Sick, Geology Department, Laboratory of Geosciences, Geomatics, and Environment,
B.P. 7955, Sidi Othmane, Casablanca (Morocco)
GeoBioStratData.Consulting, 385 Route du Mas Rillier, 69140
Rillieux-la-Pape (France)
Published online in final form (pdf) on January 14, 2026
DOI 10.2110/carnets.2026.2602
![]()
[Editor:
Alberto Collareta; language editor: Robert
W. Scott;
technical editor: Bruno R.C. Granier]
![]()
Because of the convergence of Africa and Eurasia and the opening of the Atlantic Ocean, the Gulf of
Hammamet, which makes up the southwestern portion of the Mediterranean, experienced significant relative sea-level fluctuations and eustatic variations during the late Cenozoic.
The sedimentary successions deposited between the Late Miocene and the earliest Pleistocene in the Gulf of Hammamet preserve precious environmental and climatic information. The present paper focuses on the latest Messinian to Gelasian deposits of the Jiriba-1 borehole, which is located off the coast of Tunisia. Here, previous datings obtained through planktonic foraminiferal biostratigraphy are calibrated chronologically on a high-resolution scale. Palynological analysis based on the spores and pollen content of the borehole allows for reconstructing the environmental and vegetational evolution, as well as establishing a precise climatostratigraphic framework for the study area. The latest Messinian to Gelasian palynological record is comprised of abundant continental (pollen and spores) and marine (dinoflagellate cysts) inputs. Distality markers (halophytes, D/S, Pinus, and dinoflagellate cysts) indicate that the Gulf of Hammamet was an epicontinental setting that evolved from a proximal to a distal neritic environment.
The Lygeum, Artemisia, and S/T ratio curves enable climatostratigraphic correlations with northwestern Europe, the northwestern Mediterranean, and the Black Sea. Furthermore, the climatic evolution depicted by the steppe elements (Artemisia and Ephedra) from the Jiriba-1 borehole shows parallelisms with the δ18O isotope curve.
The vegetation cover was open, dominated by herbaceous plants that inhabited the lower altitudes, and developed into mixed forest-plant formations at mid-altitudes. Between these two formations, a group of trees made up of Mediterranean species was present.
Repetitive changes in vegetation as well as relative sea-level fluctuations in the Gulf of Hammamet suggest that these cyclical variations resulted from the influence of astronomical forcing.
• climate;
• vegetation;
• climatostratigraphy;
• southwestern Mediterranean;
• Gulf of Hammamet;
• Tunisia
Berry N., Bachiri Taoufiq N., Barhoun N., Suc J.-P., Essamoud R., Targhi S., Bahaj H., Rahmouna J. & Popescu S.-M. (2026).- Climate and vegetation changes in the southwestern Mediterranean during the Plio-Pleistocene: Palynological and climatostratigraphic record of the Gulf of Hammamet (NE Tunisia).- Carnets Geol., Madrid, vol. 26, no. 2, p. 21-49. DOI: 10.2110/carnets.2026.2602
Changements climatiques et de la végétation dans le sud-ouest de la Méditerranée au cours du Plio-Pléistocčne : Enregistrements palynologiques et climatostratigraphiques dans le golfe de Hammamet (NE Tunisie).- En raison de la convergence entre les plaques africaine et eurasienne ainsi que de l'ouverture de l'océan
Atlantique, le golfe de Hammamet, qui constitue la partie sud-ouest du bassin méditerranéen, a connu d'importantes fluctuations du niveau relatif de la mer ainsi que des variations eustatiques au cours du Cénozoďque
supérieur.
Les successions sédimentaires déposées entre le Miocčne supérieur et le Pléistocčne inférieur dans le golfe de Hammamet renferment de précieuses informations environnementales et
climatiques. Notre étude se concentre sur les dépôts du Messinien terminal au
Gélasien reconnus dans le forage Jiriba-1 situé au large des côtes
tunisiennes. Dans ce cadre, les datations antérieures basées sur la biostratigraphie des foraminifčres planctoniques sont ici recalibrées sur une échelle chronostratigraphique ŕ haute
résolution.
L'analyse palynologique, basée sur le contenu en spores et pollen du forage, permet de reconstituer l'évolution de l'environnement et de la
végétation, mais aussi d'établir un cadre climatostratigraphique précis pour la zone d'étude. L'enregistrement palynologique s'étendant du Messinien terminal au Gélasien est marqué par des apports abondants d'origine continentale (pollen et spores) et marine
(kystes de dinoflagellés). Les marqueurs de distalité (halophytes, D/S, Pinus et kystes de
dinoflagellés) indiquent que le golfe de Hammamet constituait un environnement épicontinental ayant évolué
depuis un milieu néritique proximal vers un milieu néritique distal.
Les courbes de Lygeum, d'Artémisia et du rapport S/T permettent d'établir des corrélations climatostratigraphiques avec le nord-ouest de l'Europe, la Méditerranée nord-occidentale et la Mer Noire. Par
ailleurs, l'évolution climatique illustrée par les éléments steppiques (Artémisia et Ephedra) issus du forage Jiriba-1 présente des parallélismes avec la courbe isotopique δą⁸O.
La couverture végétale était ouverte, dominée par des plantes herbacées occupant les basses altitudes et évoluant vers des formations forestičres mixtes aux altitudes
moyennes. Entre ces deux formations, un groupement arboré composé d'espčces méditerranéennes
a été présent.
Les changements répétitifs de la végétation ainsi que les fluctuations relatives du niveau marin dans le golfe de Hammamet suggčrent que ces cycles résultent de l'influence
de forçages astronomiques.
• climat ;
• végétation ;
• climatostratigraphie ;
• sud-ouest de la Méditerranée ;
• golfe de Hammamet ;
• Tunisie
The Mediterranean Sea is an intercontinental basin linked to the Atlantic Ocean through the 14-km-wide Strait of Gibraltar to the west. Because of its peculiar position at the confluence between the humid temperate Central European climate and the arid North African climate with North Atlantic and tropical influences (Mehta & Yang, 2008), the western Mediterranean has undergone geological, climatic, and environmental upheavals, especially during the Neogene, a period characterized by significant geodynamic and palaeogeographic changes in this area. Processes involving subduction, continental collision, and the opening of sizable basins in a back-arc context shaped this semi-enclosed sea (Do Couto, 2014). The complicated history of the Mediterranean and the evolution of water exchange was the ultimate result of tectonic forcing combined with eustatic and climatic factors (Flecker et al., 2015).
At the beginning of the Oligocene (33.9 Ma), the NS-oriented convergence of Africa and Eurasia reduced the oceanic space between these continents along various subduction zones (Jolivet & Faccenna, 2000; Bernatchez et al., 2008; Do Couto, 2014). The Lower Miocene marked the opening of the western Mediterranean (Rehault et al., 1984; Robertson & Grasso, 1995) and the birth of the northern Betic and southern Rifian corridors (Martín Martín et al., 2014; Flecker et al., 2015; Capella et al., 2018; Krijgsman et al., 2018). This resulted from the westward displacement of the Alboran domain and the continued convergence of the African and Iberian plates (Jolivet & Faccenna, 2000; Faccenna et al., 2004; Do Couto, 2014). These two corridors enabled the exchange of water between the North Atlantic and the Mediterranean Sea during the Late Miocene (Bachiri Taoufiq, 2000; Barhoun & Bachiri Taoufiq, 2008; Flecker et al., 2015; Krijgsman et al., 2018) and the opening of the Aegean Sea during the Tortonian (Steininger et al., 1988). During the subsequent Messinian, significant tectonic activity occurred because of a N-S-oriented compressional regime (Serpelloni et al., 2007; Bernatchez et al., 2008; Do Couto, 2014). This tectonic instability, combined with geodynamic processes and eustatic fluctuations, led to the successive closure of the Betic (7.2 Ma) and Rifian corridors (7.35-7.25 Ma in the north, 7.1-6.9 Ma in the south), and eventually to the constriction of the proto-Strait of Gibraltar (Do Couto, 2014; Martín Martín et al., 2014; Flecker et al., 2015; Capella et al., 2018; Krijgsman et al., 2018; Spakman et al., 2018; Targhi et al., 2021). As a consequence of this, the latter became the only open gateway to the Atlantic during the Messinian, and probably delivered salty water to the Mediterranean during all successive stages of the Messinian Salinity Crisis (5.97-5.333 Ma) (Hsü et al., 1973; Krijgsman et al., 1999; Jolivet et al., 2006; Clauzon et al., 2015; Suc & Frizon de Lamotte, 2019; Suc et al., 2020). This phenomenon constitutes the most fascinating oceanographic event recorded in recent Mediterranean history (Cornée et al., 2016) - one that led the Mediterranean to experience major salinity fluctuations and hypersaline conditions (Hsü et al., 1973; Krijgsman et al., 1999; Roveri et al., 2014; Dijk et al., 2023) as testified by the deposition of massive evaporitic successions in the peripheral and central Mediterranean basins over a very short time span (Rouchy, 1982; Rouchy & Caruso, 2006; Krijgsman & Meijer, 2008; Roveri et al., 2014).
The reflooding of the Mediterranean basin through the reopening of the Strait of Gibraltar has been estimated to have begun around 5.46 Ma, before the onset of the Zanclean. It resulted in the restoration of fully marine conditions in most of the depressions bordering the Alboran Sea, as well as in a sudden, generalized rise of the Mediterranean sea-level (Steininger & Rögl, 1984; Blanc, 2002; Loget & Driessche, 2006; García-Castellanos et al., 2009; Estrada et al., 2011; Bache et al., 2012; Suc & Frizon de Lamotte, 2019; Suc et al., 2020).
Situated between Africa and Eurasia, the Mediterranean region was colonized by vegetation combining tropical, northern, and native flora (Feddi et al., 2011). The rich fossil pollen record from this area provides a detailed history of plant ecosystems, and indicates tropical to subtropical climatic conditions from the early Neogene (23.04 Ma) to the latest Pliocene (2.6 Ma) (Suc, 1984, 1989; Bessedik, 1985; Suc & Popescu, 2005; Jiménez-Moreno et al., 2009, 2013, 2015; Suc et al., 2018b; Bertini & Combourieu-Nebout, 2023; Bertini et al., 2025). In the northern Mediterranean, over the past 23 million years, thermophilous and hygrophilous taxa declined, which was accompanied by a transition from dense forests to open landscapes. Avicennia mangroves first migrated southward before disappearing and being replaced by swamps that also vanished subsequently. Subtropical evergreen forests were successively replaced by sclerophyllous Mediterranean vegetation and then by Artemisia steppes, alternating with mesophilous forests during glacial-interglacial cycles. In the southern Mediterranean, open subdesertic associations prevailed, with Avicennia persisting until the Early Pliocene. Mediterranean sclerophyllous plants, which first appeared in the Early Miocene, expanded during the Neogene in response to climatic changes, with subsequent variations being influenced by temperature and seasonal instability (Suc et al., 2018b).
Pollen analysis shows that the Mediterranean changing climate can be subdivided into three main phases covering the past 23 million years (Suc et al., 2018b). After a phase of relatively stable, warm conditions with limited thermal seasonality, ranging from the Miocene to the early Late Pliocene (23 - 3.37 Ma), the 3.37-2.6 Ma time span represents a transitional phase during which a Mediterranean-type seasonal rhythm was established. Around 2.6 Ma, floristic diversity declined, and plant communities underwent major transformations as a consequence of the Intensification of Northern Hemisphere glaciation (INHG), which was initiated around 2.7 Ma (Mudelsee & Raymo, 2005; Kaboth-Bahr et al., 2021; McClymont et al., 2023; Bertini et al., 2025). This led to widespread xericity and cold temperatures, with highly contrasting seasons, while interglacial periods were warmer and wetter (Popescu, 2001; Suc et al., 2018b). In addition to these long-term changes, Zachariasse et al. (1989), Hilgen (1990, 1991), Langereis and Hilgen (1991), and Laan et al. (2006) have demonstrated that, in Sicily and Morocco, the variability of orbital parameters strongly controlled marine sedimentation during the Early Pliocene. In the Mediterranean region, cyclical changes in vegetation during the Pliocene were mainly due to precession (Popescu, 2001; Popescu et al., 2010; Jiménez-Moreno et al., 2013).
The Gulf of Hammamet has been the focus of intense oil exploration activity in recent years. It is part of the Southeast Mediterranean region and comprises a sizable sedimentary reservoir with precious geological and climatic archives. It is home to several basins, which mostly accommodated deposition during the Pliocene and Quaternary (Ben Brahim et al., 2013). The sedimentary fill of these basins represents a valuable paleoenvironmental archive for characterizing and understanding the last few million years of climate change in the Mediterranean Basin.
Data on vegetation, climate, and their evolution between the latest Messinian to the Gelasian are scarce in Tunisia and the surrounding countries (Algeria and Morocco). The only available studies are those of Suc et al. (1995) and Fauquette et al. (1999) on Oued et Tellil in Tunisia and Habibas-1 in Algeria, as well as those of Feddi et al. (2011) and Warny (1999) concerning Nador-1 and Bou Regreg in Morocco. To fill this gap, we conducted a palynological study based on pollen and spore analysis of the Jiriba-1 borehole, located on the eastern coast of Tunisia.
The goal of the present work is threefold. Firstly, it aims at reconstructing the evolution of vegetation and climate in the Gulf of Hammamet between the latest Messinian and Gelasian within a precise chronostratigraphic framework. Secondly, it focuses on reconstructing the continental and marine environments, and placing them in the overall context of climate change in the Mediterranean region. Thirdly, it aims at investigating how vegetation along the southwestern Mediterranean coast reacted to the forcing of orbital parameters.
Northern Tunisia is bordered by the Mediterranean Sea, and its coastline is deeply indented by numerous embayments, most notably the gulfs of Tunis, Hammamet, and Gabes (Tayech,
1984) (Fig. 1 ![]() ).
).
Our study area, the Gulf of Hammamet, is located in the offshore zone of northeastern Tunisia, between two major geological domains: the Atlas domain to the west and the Pelagian domain to the east (Ben Brahim et al.,
2013). It is bounded by the Cap Bon Peninsula to the north, the Sicily Channel to the northeast, the Sahel province to the west, and the Mahdia-Kerkennah uplift to the south (Boccaletti et al.,
1987; Gardiner et al.,
1995; Torelli et al., 1995; Catalano et al.,
1996; Tavarnelli et al.,
2004; Ben Brahim et al.,
2013). Its surface area covers approximately 20,000 km˛ (Ben Brahim et al.,
2013)
(Fig. 1 ![]() ).
).
The northeastern region of Tunisia is characterized by a significant accumulation of Neogene deposits resulting from continuous subsidence during the Oligocene-Miocene to the Pliocene (Benzakour, 1992; Dlala, 1994, 1995; Kacem, 2004; Mejri, 2012). Resulting from the erosion of the surrounding reliefs, these deposits filled the subsiding basins, which are segmented by horsts and grabens (Mejri, 2012). The Miocene successions in the Gulf of Hammamet can reach up to 2,500 m in thickness and are mainly composed of clayey, sandstone units, along with gypsum (Mejri, 2012). Phytogenic deposits interbedded within these mostly siliciclastic successions indicate a paralic setting (Ben Moktar & Mannaď-Tayech, 2012).
The Messinian deposits of northeastern Tunisia - including the Kechabta, Oued Belkhedim, and Oued Et Tebala formations - are unconformably overlain by the Lower Pliocene Raf-Raf Formation, which is mainly composed of marls and clays. This unit is further overlain by the Upper Pliocene Porto Farina Formation, which is dominated by sandstone deposits. The succession formed by the Raf-Raf and Porto Farina formations contains terrestrial mammal fossils assigned to the Early Villafranchian (Laffitte & Dumon, 1948; Bizon et al., 1980; Besčme & Lajmi, 1981; Mejri, 2012).
Overall, the Mio-Pliocene deposits constitute a regressive megasequence bounded at its base by a discontinuity marking the Tortonian-Messinian transition, and at the top by a second discontinuity associated with the Pliocene transgression (Besčme & Kamoun, 1988; Gaaloul & Razgallah, 2008; Frigui et al., 2016).
The sedimentary settings of northeastern Tunisia evolved from a shallow marine carbonate platform characterized by a diverse fauna to a lagoonal setting marked by evaporitic sedimentation, and eventually to a shallow-water, restricted environment. This palaeoenvironmental succession extends in a nearly continuous fashion between the Messinian and the Quaternary (Temani & Gaaloul, 2007; Gaaloul & Razgallah, 2008).
The tectonic framework of the Gulf of Hammamet reflects the dynamic convergence between the African and Eurasian plates, as well as the opening of the Atlantic Ocean (Chihi & Philip, 1999; Mannaď-Tayech, 2006; Ben Brahim et al., 2013). At different times, these processes have induced both compressive and extensive tectonic regimes, leading to a variety of structures and tectonic inversions of sedimentary basins (Ben Ayed et al., 1983; Burollet, 1991; Bédir et al., 1992; Bensalem, 1992; Piquet et al., 2002; Ben Brahim et al., 2013; Abidi et al., 2015; Saint-Martin et al., 2023).
The study area is marked by a major north-south-oriented structure, crossing the entire Atlas domain of central Tunisia (Bouaziz et al.,
2002; Ben Brahim et al.,
2013)
(Fig. 1 ![]() ). To the east of this north-south axis, the Pelagian domain is subdivided into two distinct zones: on the one hand, the continental Sahel domain, where upper Cenozoic sediments are buried beneath a folded zone affected by major NW-SE- and E-W-trending normal faults; on the other hand, the offshore Pelagian Sea, which is characterized by a set of shallow platforms and intermediate basins controlled by a complex fault network (Haller,
1983; Bédir, 1995; Argnani & Torelli,
2001; Ben Brahim et al.,
2013).
). To the east of this north-south axis, the Pelagian domain is subdivided into two distinct zones: on the one hand, the continental Sahel domain, where upper Cenozoic sediments are buried beneath a folded zone affected by major NW-SE- and E-W-trending normal faults; on the other hand, the offshore Pelagian Sea, which is characterized by a set of shallow platforms and intermediate basins controlled by a complex fault network (Haller,
1983; Bédir, 1995; Argnani & Torelli,
2001; Ben Brahim et al.,
2013).
During the Middle Miocene (Langhian), the Hammamet Basin underwent a very strong subsidence with a subsidence rate reaching 150 m/Ma (Patriat et al., 2003; Ben Brahim et al., 2013). This conspicuous subsidence was synchronous with the emplacement of thrust sheets in the Tellian domain (Rouvier, 1977). During the Late Miocene, the northeastern region of Tunisia experienced a tectonic peak marked by the combined activity of Atlasic and Alpine phases. This geodynamic evolution led to the formation of the Alpine chains, such as the Kroumirie in the north and northwest, and the antiform of Jebel Abderrahmane in the northeast. It also contributed to the structuring of the Tunisian Ridge and the individualization of several microbasins, which were filled with molassic and fluviodeltaic siliciclastic deposits organized into sedimentary sequences (Mannaď-Tayech, 2006; Ben Moktar & Mannaď-Tayech, 2012).
In Pliocene and Pleistocene times, the Gulf of Hammamet and the nearby Kuriate Islands were affected by significant subsidence associated with extensional tectonics linked to the active Siculo-Tunisian rift, which resulted in the development of horst and graben structures since the Oligocene and Miocene (Haller, 1983; Burollet, 1991; Benzakour, 1992; Bédir, 1995; Bédir et al., 1996; Ben Brahim et al., 2013). Subsequently, the post-Villafranchian compressive phase triggered the formation of the Jiriba trough by reactivating pre-existing normal faults as reverse faults (Ben Brahim et al., 2013).
This subsiding basin is home to significant Cretaceous and Miocene-Pliocene reservoirs, which provide favorable conditions for hydrocarbon accumulation due to their structural geometries (Abidi et al., 2015).
|
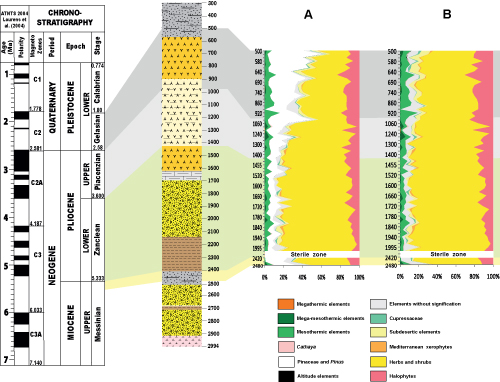
Figure 1:
A)
Simplified geological map of northeastern Tunisia (Benzakour,
1992); B) Location map of the studied region showing the main structural
elements. |
The Jiriba-1 borehole, located in the Gulf of Hammamet (Fig. 1 ![]() ) approximately 18 km offshore from the eastern Tunisian coast
(36°12'N,
10°40'E), was drilled to a total depth of 2885 m. Samples were collected in 1982 by
elf-Aquitaine Tunisie and Shell as part of an exploration program targeting Upper Miocene (Messinian) to Quaternary deposits (Bismuth & Benzarti,
1982; Londeix et al., 1999).
) approximately 18 km offshore from the eastern Tunisian coast
(36°12'N,
10°40'E), was drilled to a total depth of 2885 m. Samples were collected in 1982 by
elf-Aquitaine Tunisie and Shell as part of an exploration program targeting Upper Miocene (Messinian) to Quaternary deposits (Bismuth & Benzarti,
1982; Londeix et al., 1999).
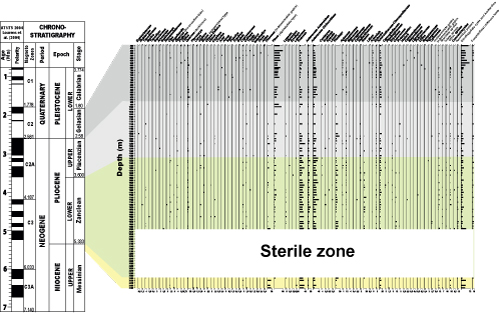
Palynological slides from the Jiriba-1 borehole are stored at the Laboratory of Geosciences, Geomatics, and Environment, Geology Department, Faculty of Sciences Ben M'Sick, Hassan II University of Casablanca (Morocco). The borehole spans between the Messinian and the base of the Gelasian and features a reasonably continuous Pliocene succession (Fig.
2 ![]() ). The deposits were thoroughly described by Bismuth and Benzarti
(1982), Benzakour (1992), and Londeix et al.
(1999), who distinguished two main formations
(Fig.
2
). The deposits were thoroughly described by Bismuth and Benzarti
(1982), Benzakour (1992), and Londeix et al.
(1999), who distinguished two main formations
(Fig.
2 ![]() ):
):
Sampling for palynology was carried out every 20 m between depths of 2994 m and 300 m. The samples analyzed in this study are located between depths of 2480 and 500 m.
Bismuth and Benzarti (1982) conducted the first micropalaeontological study of the Jiriba-1 borehole. It focused on benthic and planktonic foraminifera as well as on ostracods. Based on the vertical distribution of planktonic foraminiferal marker species, Benzakour (1992) and Londeix et al. (1999) proposed a stratigraphic subdivision of the succession. The bioevents considered in this study correspond to the first and last appearances of the following taxa: Sphaeroidinellopsis, Globorotalia margaritae, Globorotalia puncticulata, Globorotalia crassaformis, and Globorotalia inflata. These events are presented sequentially, from top to bottom (Table 1).
Table 1: Main Planktonic Foraminiferal Bioevents in the Jiriba-1 Borehole.
| Stage | Planktonic foraminifera bioevents | Depth (m) | Age (Ma) |
| Gelasian | First Occurrence (FO) of Globorotalia inflata | 620 m | 2.09 Ma |
| Last Regular Occurrence (LRO) of Globorotalia crassaformis | 880 m | 2.13 Ma | |
| Piacenzian | Last Occurrence (LO) of Sphaeroidinellopsis | 1280 m | 3.19 Ma |
| Zanclean | Last Occurrence (LO) of Globorotalia puncticulata | 1420 m | 3.57 Ma |
| Last Common Occurrence (LCO) of Globorotalia margaritae | 1620 m | 3.98 Ma | |
| First Occurrence (FO) of Globorotalia puncticulata | 1960 m | 4.52 Ma | |
| First Common Occurrence (FCO) of Globorotalia margaritae | 2320 m | 5.08 Ma | |
| Zanclean Base | Acme Base (AB) of Sphaeroidinellopsis | 2400 m | 5.30 Ma |
In the present work, these bioevents were calibrated with the Mediterranean stratigraphic scale (Lourens et al.,
2004; Iaccarino et al.,
2007; Lirer et al.,
2019), which allowed us to recognize the uppermost Messinian, the Pliocene (Zanclean and Piacenzian), and the Gelasian
(Fig.
2 ![]() ). According to the most recent version of the International Commission on Stratigraphy (ICS), the Gelasian stage is regarded herein as part of the Lower Pleistocene.
). According to the most recent version of the International Commission on Stratigraphy (ICS), the Gelasian stage is regarded herein as part of the Lower Pleistocene.
The uppermost Messinian (5.5 Ma to 5.333 Ma) corresponds to depths between 2480 and 2400 m, directly below the AB Sphaeroidinellopsis (Sphaeroidinellopsis Acme Base) bioevent, corresponding to 5.333 Ma (Lourens et al.,
2004), which marks the restoration of open-marine conditions in the Mediterranean Basin following the Messinian Salinity Crisis
(Fig.
2 ![]() ).
).
The Zanclean (Lower Pliocene) (5.333 Ma to 3.600 Ma) extends from 2400 to 1420 m in depth, according to previously cited studies. Palynologically, the 2400-2000 m interval is barren, whereas only the samples between 2000 and 1420 m are rich in terrestrial and marine palynomorphs. This interval corresponds to the upper part of the Zanclean, specifically to the MPL3 zone and MPL4a subzone
(Fig.
2 ![]() ).
).
The MPL3 Zone (2000 to 1700 m) corresponds to the interval between the FO of Globorotalia puncticulata and the LCO of Globorotalia margaritae, from 4.52 Ma to 3.98 Ma (Lourens et al.,
2004). The interval between the LCO of Globorotalia margaritae and the LO of Globorotalia puncticulata, from 3.98 Ma to 3.57 Ma, coincides with subzone MPL4a (1700 to 1420 m) (Lourens et al.,
2004)
(Fig.
2 ![]() ).
).
The Zanclean-Piacenzian boundary (3.600 Ma) is located at a depth of 1420 m and coincides with the LO of Globorotalia puncticulata (Lourens et al.,
2004; Lirer et al.,
2019)
(Fig.
2 ![]() ).
).
The Piacenzian (Upper Pliocene) (3.600 Ma to 2.58 Ma) extends between depths of 1420 m and 960 m (Fig.
2 ![]() ).
).
The Piacenzian-Gelasian boundary (2.58 Ma; Lourens et al.,
2004) is marked by the LCO of Globorotalia bononiensis (Lourens et al.,
2004). However, according to previous studies, this species was not recorded in the Jiriba-1 borehole. In turn, the LRO of Globorotalia crassaformis, dated at 2.134 Ma, was identified at a depth of approximately 880 m. Therefore, the base of the Gelasian would be slightly deeper, between 1060 m and 880 m, most likely around 960 m, based on the attempted correlation of this borehole with the reference Rossello section (Fig.
2 ![]() ).
).
The Gelasian (lowermost Pleistocene) (2.58-1.80 Ma) is characterized by two events, namely, the FCO of Neogloboquadrina pachyderma with sinistral coiling and the influx of Globorotalia crassaformis (Lourens et al.,
2004). In the studied borehole, only one bioevent was observed at around 620 m, corresponding to the FO Globorotalia inflata, while the appearance of Globorotalia truncatulinoides was not recorded. Therefore, the sediments between 960 m and 500 m are of Gelasian age, probably corresponding to subzone MPL6a. The latter spans between the FO of Globorotalia inflata and the FO of Globorotalia truncatulinoides (Lourens et al.,
2004; Iaccarino et al.,
2007)
(Fig.
2 ![]() ).
).
Remarks: The biostratigraphic subdivisions adopted herein for the Jiriba-1 Borehole generally correlate with the major climatic phases thereof (see Discussion below). Indeed, the MPL3 zone corresponds to the Pollen sub-zone P Ib of Suc and the Brunssumian B of Zagwijn (from 4.75 Ma to 4.0 Ma; 2000 to 1700 m), while the MPL4a zone coincides with the pollen sub-zone P Ic of Suc and the Brunssumian C of Zagwijn (from 4.0 Ma to 3.600 Ma; 1700 to 1420 m). The Piacenzian (3.600 Ma to 2.58 Ma) is consistent with the P II zone of Suc and the Reuverian of Zagwijn (1420 to 960 m). The Gelasian (2.58 Ma to 1.80 Ma; 960 to 500 m) matches the P III zone of Suc and the Praetiglian of Zagwijn (Zagwijn, 1960; Suc & Zagwijn, 1983; Suc, 1984; Suc et al., 1995).
|
Figure 2:
Lithostratigraphy and biostratigraphy of the Jiriba-1 borehole. |
3.4.1. Palynological technique
This method aims at progressively eliminating all mineral and organic constituents, thereby concentrating the palynomorphs. To achieve this, samples from the Jiriba-1 borehole were prepared using the classical palynological technique adopted by Popescu (2001), following the steps below:
3.4.2. Bioclimatic markers of continental environments
The mean annual temperature (MAT) can be inferred from ecological data derived from current plant assemblages (Nix, 1982), including megatherm, mega-mesotherm, mesotherm, and microtherm elements (Suc et al., 2018b). Humidity can be estimated using the Poaceae/Compositae ratio (Cour & Duzer, 1978).
These correspond to the megatherm plants from equatorial and tropical forests (MAT>24°C). The group includes Euphorbiaceae, Rubiaceae, Hamamelidaceae, and Malvaceae.
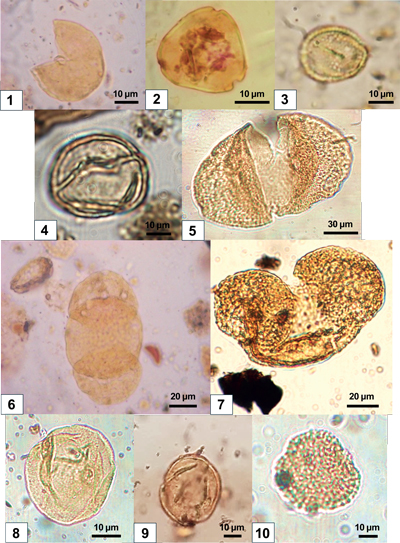
These correspond to plants living in subtropical forests (24°C>MAT>20°C). This group includes Taxodiaceae, Myrica, Engelhardia
(Pl. 1 ![]() , fig. 2), Sapotaceae, Arecaceae, Sapindaceae, Hedera, and
Magnoliaceae.
, fig. 2), Sapotaceae, Arecaceae, Sapindaceae, Hedera, and
Magnoliaceae.
These correspond to mesotherm plants typical of warm-temperate forests (20°C>MAT>14°C). This group includes Carya, Quercus (deciduous), Ulmus-Zelkova, Ulmus, Buxus sempervirens
type
(Pl. 1 ![]() , fig.
3), Carpinus, Juglans, Pterocarya, Corylus, Acer, Alnus, Salix, Populus, Fraxinus, Betula, Ericaceae, and Ligustrum.
, fig.
3), Carpinus, Juglans, Pterocarya, Corylus, Acer, Alnus, Salix, Populus, Fraxinus, Betula, Ericaceae, and Ligustrum.
These correspond to plants growing in cool temperate forests at mid-altitudes, including Cathaya
(Pl. 1 ![]() , fig.
5), Tsuga, and Cedrus (14°C>MAT>12°C).
, fig.
5), Tsuga, and Cedrus (14°C>MAT>12°C).
These are characteristic of high-altitude boreal conifer forests such as Abies and Picea (MAT<12°C).
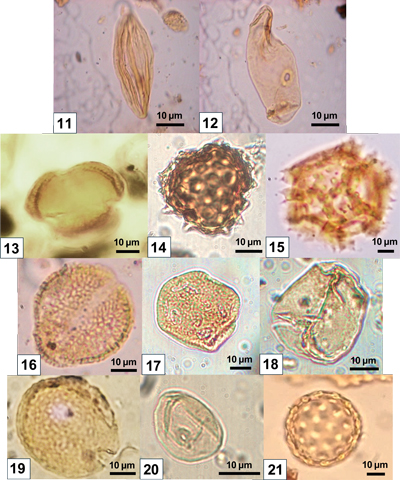
This curve represents the ratio between Poaceae
(Pl. 2 ![]() , fig.
20) and Compositae, and reflects variations in humidity, particularly in xeric environments (Cour & Duzer,
1978). Dry conditions favor the development of Compositae (ratio <20, when annual precipitation is less than 500 mm), whereas humid conditions favor the development of Poaceae (ratio >20, when annual precipitation exceeds 500 mm). This contrasts with the genus Lygeum
(Pl. 2
, fig.
20) and Compositae, and reflects variations in humidity, particularly in xeric environments (Cour & Duzer,
1978). Dry conditions favor the development of Compositae (ratio <20, when annual precipitation is less than 500 mm), whereas humid conditions favor the development of Poaceae (ratio >20, when annual precipitation exceeds 500 mm). This contrasts with the genus Lygeum
(Pl. 2 ![]() , fig.
12), which currently occurs under arid climatic conditions characterized by a mean annual temperature between 16 and 21°C and a mean annual precipitation of approximately 150-400 mm (Fauquette et al.,
1998b).
, fig.
12), which currently occurs under arid climatic conditions characterized by a mean annual temperature between 16 and 21°C and a mean annual precipitation of approximately 150-400 mm (Fauquette et al.,
1998b).
3.4.3. Marine environmental index
These indexes provide an overview of the sea-level variations that affected the Gulf of Hammamet during the latest Miocene, Pliocene, and the earliest stage of the Pleistocene. The following indicators were applied:
Due to their pollen morphology, Pinaceae (balloon grains)
(Pl. 1 ![]() ,
figs. 6-7) are easily transported by air and water, which enables a wide dispersal. In contrast, halophytes are reliable indicators of coastal environments. Generally speaking, Pinaceae are proportionally more abundant in distal than proximal areas (Heusser & Balsam,
1977; Suc & Drivaliari,
1991).
,
figs. 6-7) are easily transported by air and water, which enables a wide dispersal. In contrast, halophytes are reliable indicators of coastal environments. Generally speaking, Pinaceae are proportionally more abundant in distal than proximal areas (Heusser & Balsam,
1977; Suc & Drivaliari,
1991).
The ratio between the number of dinocysts (D) and the number of spores + pollen (S) is used to assess the marine influence (relative abundance of dinocysts) versus continental influence (pollen and spores) (Warny, 1999). This ratio allows for the recognition of transgressive-regressive trends, thus providing an excellent indicator of sea-level dynamics in marine depositional settings.
The detailed palynological analysis of the Jiriba-1 borehole reveals a significant richness of pollen and dinoflagellate cysts. The latter have been previously studied in detail by Benzakour (1992) and Londeix et al. (1999). Sediments at the base of the Zanclean were found to be devoid of palynomorphs, possibly due to oxidation of the organic matter (Benzakour, 1992).
This region has a diverse and relatively abundant pollen flora. A total of 15,671 palynomorphs were counted across 63 samples, which led to the identification of 75 pollen taxa. The detailed and synthetic diagrams
(Figs. 3 ![]() - 4
- 4 ![]() ) reflect a rather homogeneous floristic composition from the latest Messinian to the Gelasian.
) reflect a rather homogeneous floristic composition from the latest Messinian to the Gelasian.
The genus Pinus (2 to 53%) dominates the pollen spectra throughout the section. Notably, Pinus declines at the top of the Lower Pliocene and in the basal part of the Upper Pliocene (2000 m to 1100 m), with slight increases at sample depths of 1720 m, 1660 m, 1280 m, 1260 m, and 1200 m
(Figs. 3 ![]() , 4.A
, 4.A ![]() ).
).
Altitude elements (microtherm elements) ranging from 0 to 0.35% appear only at the top of the Upper Pliocene, with Abies (0.35%) and Picea (0.34%)
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
Cathaya (0 to 1.53%) and Cupressaceae (0.7 to 2.5%) occur sporadically. Cathaya appears discontinuously, alternating with the histograms of steppe elements (Artemisia and Ephedra)
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
Megatherm elements occur very rarely, with frequencies ranging from 0 to 0.68%
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
Elements of the mega-mesotherm group (Arecaceae, Cupressaceae, Engelhardia, Magnoliaceae, Myrica, Sapindaceae, Sapotaceae, and Sciadopitys) are present along the profile at low frequencies, ranging from 0 to 4.89%
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
Elements of the mesotherm group (ranging from 0.43 to 16.23%) include, in order of abundance, Quercus (deciduous) (2 to
16%)
(Pl. 1 ![]() , fig.
4), Buxus sempervirens type (0 to 3.38%), and Juglans (0 to 1%). Other elements (Acer, Hedera, Alnus, Betula, Carpinus, Corylus, Ericaceae, Carya, Pterocarya, Fraxinus, Ligustrum, Populus, Salix, and Ulmus-Zelkova) do not exceed 0.7%
(Figs. 3
, fig.
4), Buxus sempervirens type (0 to 3.38%), and Juglans (0 to 1%). Other elements (Acer, Hedera, Alnus, Betula, Carpinus, Corylus, Ericaceae, Carya, Pterocarya, Fraxinus, Ligustrum, Populus, Salix, and Ulmus-Zelkova) do not exceed 0.7%
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
The Mediterranean group (0 to 3.40%) is characterized by the presence of Quercus ilex-type
(Pl. 1 ![]() , fig.
9) (1 to 3%), Olea
(Pl. 1
, fig.
9) (1 to 3%), Olea
(Pl. 1 ![]() , fig.
10) (0 to 1.1%), Phillyrea (0 to 0.8%), and Cistus
(Pl. 1
, fig.
10) (0 to 1.1%), Phillyrea (0 to 0.8%), and Cistus
(Pl. 1 ![]() , fig.
8) (0 to 0.6%)
(Figs. 3
, fig.
8) (0 to 0.6%)
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
The herbaceous plants are very abundant, representing more than 83% of the pollen sum
(Fig. 4.B ![]() ). Within this group, the Compositae (Asteraceae) Asteroideae
(Pl. 2
). Within this group, the Compositae (Asteraceae) Asteroideae
(Pl. 2 ![]() , fig.
14) and Compositae (Asteraceae) Cichorioideae
(Pl. 2
, fig.
14) and Compositae (Asteraceae) Cichorioideae
(Pl. 2 ![]() , fig.
15) dominate the pollen spectra, with frequencies between 3 and 40%
(Fig. 3
, fig.
15) dominate the pollen spectra, with frequencies between 3 and 40%
(Fig. 3 ![]() ). Poaceae are between 4 and 37%, and Plantago pollen
(Pl. 2
). Poaceae are between 4 and 37%, and Plantago pollen
(Pl. 2 ![]() , fig.
19) reaches values up to 25%
(Fig. 3
, fig.
19) reaches values up to 25%
(Fig. 3 ![]() ). Note that where Poaceae pollen frequencies decrease (e.g., between 1860 m and 1800 m), those of Compositae (Asteraceae) increase; conversely, where Poaceae frequencies increase (e.g., between 1660 m and 1580 m), those of Compositae (Asteraceae) decrease
(Fig. 3
). Note that where Poaceae pollen frequencies decrease (e.g., between 1860 m and 1800 m), those of Compositae (Asteraceae) increase; conversely, where Poaceae frequencies increase (e.g., between 1660 m and 1580 m), those of Compositae (Asteraceae) decrease
(Fig. 3 ![]() ).
).
The Poaceae/Compositae ratio is close to or below 2 throughout the borehole, except for two horizons
(2480 m and 1300 m) where it exceeds 2
(Fig. 6 ![]() ).
).
The aridity of the environment is indicated by the presence of subdesertic elements (Lygeum; 0-5.58%, excluding Pinaceae) and steppe elements
[Artemisia
(Pl. 2 ![]() , fig.
13); 0-11%, excluding Pinaceae, and Ephedra
(Pl. 2
, fig.
13); 0-11%, excluding Pinaceae, and Ephedra
(Pl. 2 ![]() , fig.
11); 0-4%, excluding Pinaceae]
(Figs. 3
, fig.
11); 0-4%, excluding Pinaceae]
(Figs. 3 ![]() , 4.B
, 4.B ![]() ).
).
Halophytes (2 to 24%) are represented by
Amaranthaceae-Chenopodiaceae (Pl. 2 ![]() , fig.
21) (3 to 23%), which are dominant in the Upper Pliocene, and by Caryophyllaceae pollen
(Pl. 2
, fig.
21) (3 to 23%), which are dominant in the Upper Pliocene, and by Caryophyllaceae pollen
(Pl. 2 ![]() , fig.
17) at 1640 m (4.8%). Plumbaginaceae do not exceed 1% in the basal part of the uppermost Messinian (2480m)
(Figs. 3
, fig.
17) at 1640 m (4.8%). Plumbaginaceae do not exceed 1% in the basal part of the uppermost Messinian (2480m)
(Figs. 3 ![]() ,
4.B
,
4.B ![]() ).
).
Examination of the synthetic diagram without Pinaceae reveals that the peaks of some ecological groups show opposite variations, as observed for mega-mesotherms, Cathaya, mesotherms, subdesertic elements, and halophytes
(Fig. 4.B ![]() ).
).
The frequency of Pteridophytes spores varies between 3 and 13%, including both monolete and trilete spores (up to 9%). The presence of Concentricystes reflects fluvial input influence at three levels (1680 m, 1520 m, and 1260 m)
(Fig. 3 ![]() ).
).
Reworked pollen grains are distributed from the top of the Lower Pliocene to the Upper Pliocene (2000 m to 1140 m), with a notable increase at 1240 m
(Fig. 3 ![]() ).
).
|
Figure 3:
Detailed pollen diagram of the Jiriba-1 borehole. |
|
Figure 4:
Synthetic pollen diagrams with (A) and without (B) Pinaceae from the Jiriba-1 borehole. |
The percentage of autochthonous elements (dinoflagellate cysts, chitinous tests of foraminifera, and acritarchs) is variable throughout the borehole (Fig. 3 ![]() ).
).
During the uppermost Messinian (2480 m to 2400 m), and from the Zanclean to the early Gelasian, the D/S curve generally shows low values, ranging from 0 to 0.5, reflecting the abundance of continental inputs
(Fig. 6 ![]() ).
).
At the top of the Upper Pliocene (640 m to 500 m), the D/S curve is marked by higher ratios, exceeding 1. This reflects lower concentrations of terrestrial palynomorphs compared to dinocysts (Fig. 6 ![]() ).
).
Pinus + indeterminable Pinaceae are considered ecologically insignificant, as their pollen cannot be determined below the genus level at best. The genus Pinus includes species that span a very broad thermal range. Because of their pollen morphology, which favors wide dispersal, the abundance of these pollen grains tends to decrease in proximal marine areas and increase in distal environments (Heusser & Balsam,
1977; Suc & Drivaliari,
1991). Accordingly, the frequencies of Pinus + indeterminable Pinaceae generally follow the same trend as the D/S curve
(Fig. 6 ![]() ).
).
Halophytes, in turn, are reliable indicators of coastal environments. These indicators of distal-versus-proximal settings vary inversely. In the case of the Jiriba-1 borehole, halophyte frequencies increase as those of Pinus + indeterminable Pinaceae decrease
(Fig. 6 ![]() ).
).
The discrepancy between these distal indicators at 640 m and 1000 m may be related to tectonic activity
(Fig. 6 ![]() ).
).
After presenting a palynological synthesis of the studied borehole, this section discusses the results obtained in this study and their integration with existing data across three aspects. The first such aspect concerns the reconstruction of vegetation landscapes in the Gulf of Hammamet from the latest Messinian to the Gelasian as deduced from the pollen record. The second deals with the different climatic phases identified in the Jiriba-1 borehole based on the climatostratigraphic scale of Suc and Zagwijn (1983). The third aims to trace changes in vegetation, climate, and sea level in the Gulf of Hammamet over the same interval.
Overall, the pollen spectra from the Jiriba-1 borehole reflect a rather homogeneous floristic pattern and indicate fairly stable environmental conditions between the latest Messinian and the Gelasian
(Figs. 3 ![]() - 4
- 4 ![]() ). Vegetation was characterized by a mosaic of plant associations inherited from the Tortonian and Messinian. Consequently, in North Africa, the pollen floras of Early Pliocene age are similar to those of the subsequent Messinian (Chikhi,
1992a; Bachiri Taoufiq,
2000; Barhoun & Bachiri Taoufiq,
2008; Achalhi et al.,
2016; Targhi et al., 2023; Bahaj et al.,
2024).
). Vegetation was characterized by a mosaic of plant associations inherited from the Tortonian and Messinian. Consequently, in North Africa, the pollen floras of Early Pliocene age are similar to those of the subsequent Messinian (Chikhi,
1992a; Bachiri Taoufiq,
2000; Barhoun & Bachiri Taoufiq,
2008; Achalhi et al.,
2016; Targhi et al., 2023; Bahaj et al.,
2024).
Pollen analysis of the Jiriba-1 sediments reveals a diverse flora, comprising elements currently inhabiting the intertropical (Euphorbiaceae, Sapindaceae) and subtropical to warm-temperate regions of America and Asia (Engelhardia, Sapotaceae, Arecaceae, etc.), as well as elements that are presently found in Europe (Quercus, Ulmus, Acer, Salix, Populus, Ericaceae, Betula, Carpinus, etc.), and the Mediterranean region (Olea, Phillyrea, Quercus ilex-coccifera type, Fraxinus, etc.). Elements representative of subdesertic environments and generally xeric open vegetation (Poaceae, Lygeum, Ephedra, Artemisia, Compositae, Amaranthaceae-Chenopodiaceae, Plumbaginaceae, Plantago, Erodium, etc.) occur at high percentages
(Figs. 3 ![]() , 4.B
, 4.B ![]() ). Mid- and high-altitude elements such as Cathaya, Tsuga, Cedrus, Abies, and Picea are rare in the Miocene of North Africa but are well represented during the Pliocene and Gelasian (Suc,
1989; Suc et al., 1995, 2018b; Feddi et al.,
2011; Berry et al., 2025). Moreover, the vegetation was organized in altitudinal belts. By comparing the reconstructed vegetation of the latest Messinian-Pliocene in this region with modern vegetation in southeastern China (Wang,
1961), the Canary Islands, and the shores of the Red Sea, and based on current reference paleoenvironmental models (Suc,
1984; Jiménez-Moreno et al.,
2008, 2010; Feddi et al.,
2011; Suc et al., 2018b; Targhi et al.,
2023), the main plant formations can be identified as follows:
). Mid- and high-altitude elements such as Cathaya, Tsuga, Cedrus, Abies, and Picea are rare in the Miocene of North Africa but are well represented during the Pliocene and Gelasian (Suc,
1989; Suc et al., 1995, 2018b; Feddi et al.,
2011; Berry et al., 2025). Moreover, the vegetation was organized in altitudinal belts. By comparing the reconstructed vegetation of the latest Messinian-Pliocene in this region with modern vegetation in southeastern China (Wang,
1961), the Canary Islands, and the shores of the Red Sea, and based on current reference paleoenvironmental models (Suc,
1984; Jiménez-Moreno et al.,
2008, 2010; Feddi et al.,
2011; Suc et al., 2018b; Targhi et al.,
2023), the main plant formations can be identified as follows:
5.2.1. Climate
Pollen analysis allows for a refined reconstitution of climate evolution. The major climatic phases described by Zagwijn
(1960) in the Netherlands, Suc
(1984) in the northwestern Mediterranean, Diniz
(1984) in Portugal, and Popescu et al.
(2010) in the Black Sea and Poland have been correlated over long distances (Suc & Zagwijn,
1983; Suc et al., 1995, 2018a; Popescu, 2001)
(Fig. 7 ![]() ), and include:
), and include:
These major climatic phases correspond perfectly to the twofold division of the Pliocene and the Gelasian, respectively.
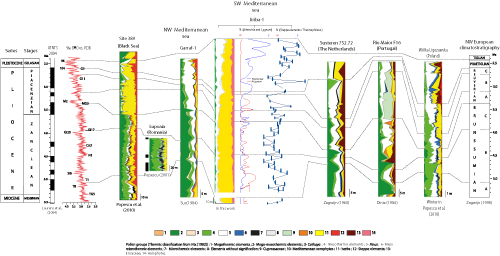
This pollen-based climatostratigraphic scale provides a benchmark for identifying the various climatic phases recorded in the Jiriba-1 borehole, corresponding to those defined by the same authors (Fig. 5 ![]() ).
).
|
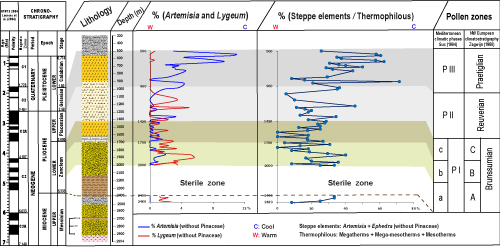
Figure 5:
Climatostratigraphic division of the Jiriba-1 borehole based on the Artemisia, Lygeum, and
steppe/thermophilous elements (S/T) curves, and their correlation with the climatostratigraphies developed in the Netherlands (Zagwijn,
1960) and in the northwestern Mediterranean region (Suc,
1984). |
The synthetic and detailed pollen diagrams from the Jiriba-1 borehole
(Figs. 3 ![]() - 4
- 4 ![]() ) reflect a repetitive change between steppe elements (Artemisia and Ephedra) and thermophiles (comprising megatherms, mega-mesotherms, and mesotherms), with no abrupt variation that facilitates the identification of major climatic phases. Consequently, it was difficult to define the exact position of any division.
) reflect a repetitive change between steppe elements (Artemisia and Ephedra) and thermophiles (comprising megatherms, mega-mesotherms, and mesotherms), with no abrupt variation that facilitates the identification of major climatic phases. Consequently, it was difficult to define the exact position of any division.
It is established that Artemisia is a good indicator of cold steppes in the western Mediterranean (Suballyova, 1997) and central continental environments (Nebout & Grazzini, 1991).
However, Lygeum is a subdesertic element indicative of a hot, dry climate. The S/T ratio curve ("Steppe elements/Thermophilous elements") is a reliable indicator of climate change (Suc et al., 2018b).
Figure 5 ![]() shows that the Lygeum and Artemisia curves are in complete opposition, with maxima of Lygeum coinciding with minima of Artemisia. Furthermore, from the base to the top of the borehole, Lygeum and Artemisia show a progressive decrease and increase, respectively. These opposing curves of Artemisia and Lygeum also correlate perfectly with the S/T ratio curve.
shows that the Lygeum and Artemisia curves are in complete opposition, with maxima of Lygeum coinciding with minima of Artemisia. Furthermore, from the base to the top of the borehole, Lygeum and Artemisia show a progressive decrease and increase, respectively. These opposing curves of Artemisia and Lygeum also correlate perfectly with the S/T ratio curve.
Thus, we used these three indicators (Artemisia, Lygeum, and S/T) to distinguish between warm and cold conditions (Fig. 5 ![]() ).
).
Analysis of these three curves reveals perfect correspondence with the climatic phases defined by Suc and Zagwijn
(1983), Suc et al. (1995), Popescu
(2001), and Suc et al. (2018a)
(Fig. 5 ![]() ).
).
Based on the comparison of the Lygeum, Artemisia, and S/T ratio curves, the following climatic phases have been identified:
This period partly coincides with the second (central and paroxysmal) stage of the Messinian Salinity Crisis, ranging from 5.60 to 5.46 Ma (Bache et al., 2012; Popescu et al., 2021; Suc et al., 2023; Dijk et al., 2023).
The latest Messinian corresponds to the absence of Artemisia. Mean frequencies of Lygeum, mega-mesotherms, and mesotherms
(Figs. 3 ![]() ,
5
,
5 ![]() ) are consistent with a warm and dry climate, similar to that recorded during the Messinian in Morocco (Bachiri Taoufiq,
2000; Bachiri Taoufiq et al.,
2008; Achalhi et al.,
2016; Targhi et al., 2021), Algeria (Chikhi,
1992a, 1992b), and southwestern Spain (Jiménez-Moreno et al.,
2013), which overall indicates that the southern Mediterranean region experienced xeric conditions climatically predisposed to desiccation (Suc et al.,
2023). This evidence suggests that global sea-level changes (global glacio-eustatism) contributed, at least in part, the onset and completion of the Messinian Salinity Crisis in the Mediterranean (Suc et al.,
2023).
) are consistent with a warm and dry climate, similar to that recorded during the Messinian in Morocco (Bachiri Taoufiq,
2000; Bachiri Taoufiq et al.,
2008; Achalhi et al.,
2016; Targhi et al., 2021), Algeria (Chikhi,
1992a, 1992b), and southwestern Spain (Jiménez-Moreno et al.,
2013), which overall indicates that the southern Mediterranean region experienced xeric conditions climatically predisposed to desiccation (Suc et al.,
2023). This evidence suggests that global sea-level changes (global glacio-eustatism) contributed, at least in part, the onset and completion of the Messinian Salinity Crisis in the Mediterranean (Suc et al.,
2023).
Sub-zone P I b: from 4.75 Ma to 4.0 Ma (2000 m to 1700 m)
Around 4.5 Ma, in the northwestern Mediterranean region of the Iberian Peninsula (Garraf-1 area, Catalonia, northeastern Spain, and Andalucía G1, offshore southeastern Spain), a cooling is indicated by the pollen data, being marked by a decrease in thermophilic trees and an increase in herbs (Suc et al.,
1995; Jiménez-Moreno et al.,
2010). This cooling is also recorded in the Dacian Basin (Popescu,
2006; Popescu et al.,
2006) and the Netherlands (Zagwijn,
1960; Suc & Zagwijn,
1983; Suc, 1984). Compared to the previous phase, this phase corresponds in the Jeriba-1 borehole to an increase in Lygeum frequency as well as to the appearance of the cold element Artemisia. The S/T curve confirms this result, indicating a trend towards climatic cooling. This variation coincides with Suc's pollen sub-zone P Ib (Suc & Zagwijn,
1983; Suc, 1984; Suc et al.,
1995) and Zagwijn's Brunssumian B
(1960), characterized by a moderately cold climate
(Fig. 5 ![]() ).
).
Sub-zone P I c: from 4.0 Ma to 3.600 Ma (1700 m to 1420 m)
This interval shows a decrease in the frequency of Artemisia and a slight decline in Lygeum, while the S/T ratio indicates warm conditions. It corresponds to the P Ic pollen sub-zone (Suc & Zagwijn,
1983; Suc, 1984; Suc et al.,
1995) and to the Brunssumian C of Zagwijn
(1960), reflecting a warm climate
(Fig. 5 ![]() ).
).
The boundary between the Zanclean and Piacenzian lies directly above the Gilbert/Gauss magnetostratigraphic transition (Berggren et al., 1985), which has recently been calibrated to 3.596 Ma (Hilgen et al., 2009). Around 3.6 Ma, the Northern Hemisphere glaciation began to intensify, culminating with the first major glacial episode, M2, at 3.3 Ma (Karas et al., 2020). This cooling is also recorded in oceanic isotope curves that appear to be generalized (Blanc et al., 1983; Thunell & Williams, 1983; Shackleton et al., 1995b; Karas et al., 2020), and may be attributed to the rapid expansion of the Antarctic ice cap and the development of glaciers in the Northern Hemisphere around 3.4 Ma, both contributing to the global temperature decline (Berger & Morel, 1992). The closure of the Panama Strait (5 Ma ago), the Betic (7.2 Ma) and Rifan corridors (7.1-6.9 Ma in its southern branch and 7.35-7.25 Ma in its northern branch), as well as the opening of the Bering Strait (at the Zanclean-Piacenzian boundary), triggered the North Atlantic thermohaline circulation, and probably also the intensification of Northern Hemisphere glaciation (Haug & Tiedemann, 1998; Bartoli et al., 2005; Berg, 2016; Capella et al., 2019). Around 3.65 Ma, the weakening of the Atlantic Meridional Overturning Circulation (AMOC), along with colder and cooler surface waters in the North Atlantic, enabled the extension of sea ice in the Arctic Ocean, which may have preconditioned the climate for the subsequent expansion of the continental ice sheet (Karas et al., 2020).
In the Nabeul-Hammamet region (northeastern Tunisia), Derbel-Damak and Zaghbib-Turki
(2002) showed that the enrichment of cold-water foraminiferal species marks the Zanclean-Piacenzian boundary. A sharp increase at the 1360 m level in the Artemisia frequency (from 0 to 3.5%) and S/T ratio (from 3.57% to 30%) indicates the first cooling event at the beginning of the Upper Pliocene in the southwestern Mediterranean
(Fig. 5 ![]() ). In the northwestern Mediterranean, the Artemisia steppes began to spread around 3 Ma, with a further significant expansion occurring around 2.6 Ma (Suc et al., 2018b). At that time, tundra-type vegetation appeared in the Netherlands (Suc & Zagwijn,
1983). In Anatolia, between 5.8 and 5.6 Ma, a pronounced expansion of Artemisia and Ephedra steppes likely correlates with the Antarctic glaciation and, subsequently, with the desiccation of the Mediterranean Sea (Suc et al., 2018b).
). In the northwestern Mediterranean, the Artemisia steppes began to spread around 3 Ma, with a further significant expansion occurring around 2.6 Ma (Suc et al., 2018b). At that time, tundra-type vegetation appeared in the Netherlands (Suc & Zagwijn,
1983). In Anatolia, between 5.8 and 5.6 Ma, a pronounced expansion of Artemisia and Ephedra steppes likely correlates with the Antarctic glaciation and, subsequently, with the desiccation of the Mediterranean Sea (Suc et al., 2018b).
According to De Schepper et al. (2009, 2013), the early Late Pliocene (3.600 to 3.0 Ma ago) was characterized by a warm global climate. However, a peak global glaciation during Marine Isotope Stage (MIS) M2 interrupted this warm phase around 3.30 Ma. Increased Pacific-to-Atlantic flow via the Central American Seaway, marked by a decline in Operculodinium centrocarpum, weakened the North Atlantic Current (NAC) between approximately 3.330 and 3.283 Ma, reducing the northward heat transport some 23-35 ka before MIS M2. The sea-level drop during this glaciation interrupted the Pacific inflow into the Atlantic, which restored the northward heat transport and ended the (MIS) M2 phase (De Schepper et al., 2009, 2013).
On the other hand, according to Suc et al. (2018b), the period from 3.37 to 2.6 Ma can be considered as representing a transitional phase corresponding to a Mediterranean-type seasonal rhythm characterized by summer droughts and cold winters. At 3.4 Ma, a temperature decrease corresponded to the cooling event that occurred in Europe (Zagwijn,
1960; Suc & Cravatte,
1982; Suc & Zagwijn,
1983; Diniz, 1984; Suc,
1984; Drivaliari et al., 1999; Popescu,
2001) linked to an increase and more significant variations in the frequency of Artemisia (Popescu,
2006; Feddi et al.,
2011). At Jeriba-1, this zone includes several cycles and is characterized by an increase in Lygeum frequency and a decline in Artemisia frequency following its abrupt rise at 1360 m
(Fig. 5 ![]() ). The percentage of Artemisia is lower than in P Ib but higher than in P Ic, while the S/T ratio shows a slight decrease compared to P Ib and an increase compared to P Ic. The climate during this phase may have been slightly warmer than in P Ib and cooler than in P Ic. This zone is consistent with pollen zone P II (Reuverian) (Suc & Zagwijn,
1983; Suc, 1984)
(Fig. 5
). The percentage of Artemisia is lower than in P Ib but higher than in P Ic, while the S/T ratio shows a slight decrease compared to P Ib and an increase compared to P Ic. The climate during this phase may have been slightly warmer than in P Ib and cooler than in P Ic. This zone is consistent with pollen zone P II (Reuverian) (Suc & Zagwijn,
1983; Suc, 1984)
(Fig. 5 ![]() ).
).
In Benzakour's (1992) study, on the same borehole, between 1420 and 960 m (in the detailed dinocyst diagram), a cooling of the sea-surface waters is also observed as an increase in the percentage of cold-water taxa (Operculodinium centrocarpum) and a decrease in warm-water taxa such as Operculodinium israelianum.
The boundary between the Piacenzian and Gelasian coincides with the Gauss/Matuyama magnetostratigraphic reversal at 2.6 Ma and also marks the onset of pronounced glacial-interglacial (G/I) fluctuations, expressed in the sawtooth pattern of the benthic δą⁸O record. Such variations, which have been documented by numerous studies (e.g., Shackleton et al.,
1995a; Lisiecki & Raymo,
2005, 2007; Bertini et al.,
2025), represent one of the major features of the Pleistocene. At the onset of glacial/interglacial (G/I) cyclicity, around 2.6 Ma, open landscapes with abundant steppe taxa became more frequent and more widespread in the northern Mediterranean (Joannin,
2007; Bertini,
2010, 2013; Feddi et al.,
2011; Combourieu-Nebout et al.,
2015). The Pliocene/Pleistocene (PII/PIII) boundary in
Figure 5 ![]() is marked by a drop in Lygeum frequency to 0% and a rise in Artemisia from 0% to 4.28%. At the same time, the S/T curve increases from 5% to 37%.
is marked by a drop in Lygeum frequency to 0% and a rise in Artemisia from 0% to 4.28%. At the same time, the S/T curve increases from 5% to 37%.
The pollen zone P III (Praetiglian) (Suc & Zagwijn, 1983; Suc, 1984) features a significant pulse of Artemisia in the northern Mediterranean - one that is associated with the onset of glacial-interglacial cycles after 2.6 Ma (Popescu, 2006; Feddi et al., 2011; Bertini, 2013).
During the Gelasian, the D/S ratio reflects a dominance of marine over continental inputs
(Fig. 6 ![]() ). This was explained by Benzakour
(1992) and Londeix et al.
(1999) by evoking the successive and progressive appearance of boreal taxa (Impagidinium pallidum, Bitectatodinium tepikiense, and Impagidinium pacificum) that depict phases of cooling in surface waters and low relative sea levels (or possibly enhanced seasonality) associated with the first Northern Hemisphere glaciations between ~2.5 and 2.1 Ma (Benzakour,
1992; Londeix et al.,
1999). This study further indicates that the interval from 960 m to 500 m is marked by a sharp rise in Artemisia frequency, reaching maximum values (11%) along this zone, reflecting an initial development of cold-climate steppe elements along with a marked decline in Lygeum frequency, which drops to 0%
(Fig. 5
). This was explained by Benzakour
(1992) and Londeix et al.
(1999) by evoking the successive and progressive appearance of boreal taxa (Impagidinium pallidum, Bitectatodinium tepikiense, and Impagidinium pacificum) that depict phases of cooling in surface waters and low relative sea levels (or possibly enhanced seasonality) associated with the first Northern Hemisphere glaciations between ~2.5 and 2.1 Ma (Benzakour,
1992; Londeix et al.,
1999). This study further indicates that the interval from 960 m to 500 m is marked by a sharp rise in Artemisia frequency, reaching maximum values (11%) along this zone, reflecting an initial development of cold-climate steppe elements along with a marked decline in Lygeum frequency, which drops to 0%
(Fig. 5 ![]() ). Simultaneously, the S/T ratio also rises, reaching a peak of 60%, indicating a corresponding decrease in thermophiles as well as cold conditions. The Artemisia and Lygeum curve peaks appear to be globally homogeneous during the Brunssumian phase, becoming more pronounced during the Praetiglian
(Fig. 5
). Simultaneously, the S/T ratio also rises, reaching a peak of 60%, indicating a corresponding decrease in thermophiles as well as cold conditions. The Artemisia and Lygeum curve peaks appear to be globally homogeneous during the Brunssumian phase, becoming more pronounced during the Praetiglian
(Fig. 5 ![]() ). This is in line with previous studies (Popescu,
2006; Popescu et al.,
2006), which highlight a more intense regional differentiation of vegetation during the Praetiglian compared to the Brunssumian.
). This is in line with previous studies (Popescu,
2006; Popescu et al.,
2006), which highlight a more intense regional differentiation of vegetation during the Praetiglian compared to the Brunssumian.
It is also concluded that in the southwestern Mediterranean (and the whole Mediterranean region for that matter) the significant vegetation interruption observed at the base of the Praetiglian (MIS 104) was marked by the development of Artemisia steppe, whereas in the Netherlands and Poland this disruption is characterized by a tundra-type environment and by Cupressaceae forest in Portugal (Popescu et al., 2006; Suc et al., 2018b).
|
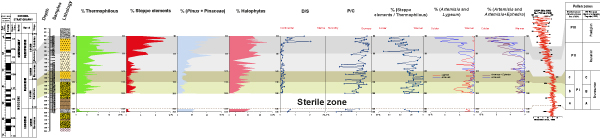
Figure 6:
Quantitative results of paleoclimatic parameters from the Jiriba-1 borehole in the Upper Miocene and Lower Pleistocene with the pollen zonation proposed by Suc and Zagwijn
(1983) and their correlation with the reference oxygen isotope curve of the ODP 846 site (Shackleton et al.,
1995b, in Suc et al., 2018b). Pollen groups: 1.
Thermophilous: Megathermic elements (Euphorbiaceae, Rubiaceae) + Mega-Mesothermic elements
(Taxodioceae, Cupressaceae, Ericaceae, Myrica, Arecaceae, Sapotaceae,
Hamamelidaceae, etc.) + Mesothermic elements (Quercus (deciduous), Carya, Juglans, Buxus sempervirens type, Ulmus-Zelkova, Corylus, Populus, etc.); 2. Steppe elements (Artemisia, Ephedra); 3. Halophytes
(Amaranthaceae Chenopodiaceae, Caryophyllaceae, Plumbaginaceae). |
5.2.2. Climatostratigraphic correlation
The Lygeum, Artemisia and S/T ratio curves correspond perfectly with the climatic phases defined by Suc and Zagwijn
(1983), Suc et al.
(1995, 2018a), and Popescu
(2001). They allow for a refined climatostratigraphic succession of the Jiriba-1 borehole (Suc's pollen sub-zones P Ib = Zagwijn's Brunssumian B; P Ic = Brunssumian C; pollen zone P II = Reuverian; pollen zone P III = Praetiglian) (Zagwijn,
1960; Suc & Zagwijn,
1983; Suc, 1984; Suc et al.,
1995)
(Fig. 5 ![]() ).
).
A correlation between these climatic phases identified here for the first time in the southwestern Mediterranean and those encountered in the northwestern Mediterranean, the Black Sea, and northwestern Europe (Suc & Cravatte,
1982; Diniz, 1984; Zagwijn,
1998; Popescu et al.,
2006) is proposed
(Fig. 7 ![]() ). Overall, this climatic subdivision is also consistent with the reference δą⁸O isotope curve (Shackleton et al.,
1995b).
). Overall, this climatic subdivision is also consistent with the reference δą⁸O isotope curve (Shackleton et al.,
1995b).
Moreover, the major phases recorded by the isotope curve are reflected across all pollen diagrams, as illustrated in
Figure 7 ![]() .
.
|
Figure 7:
Pliocene and Lower Pleistocene climatostratigraphic correlations between the synthetic pollen diagram of Jiriba-1, Site 380,
Lupoaia, Garraf-1, Susteren 752.72, Rio Maior F16 and Wólka Ligezawska; the climatostratigraphic succession of northwestern Europe (Suc et al., 2018a); and the reference oxygen isotope curve (Shackleton et al.,
1995b, in Suc et al., 2018a). |
5.3. Evolution of vegetation, climate, and sea-level variation in the Gulf of Hammamet from latest Messinian to Gelasian
5.3.1. Vegetation and climate change
Pollen diagrams from the Jiriba-1 borehole reveal a repetitive change in vegetation. Overall, the development of thermophilous elements (comprising megatherms, mega-mesotherms, mesotherms) appears to be related, on the one hand, to the decline of steppe elements (Artemisia and Ephedra) and halophytes, and on the other hand, to the increase in Pinaceae
(Fig. 6 ![]() ). It can be safely assumed that annual precipitation was well below 500 mm. These estimates are consistent with the P/C ratio (Poaceae/Compositae), which is generally below 2. We conclude that, in most sediments from the Jiriba-1 borehole, Compositae dominate over Poaceae, indicating a trend towards drought in the lowlands
(Fig. 6
). It can be safely assumed that annual precipitation was well below 500 mm. These estimates are consistent with the P/C ratio (Poaceae/Compositae), which is generally below 2. We conclude that, in most sediments from the Jiriba-1 borehole, Compositae dominate over Poaceae, indicating a trend towards drought in the lowlands
(Fig. 6 ![]() ). We also conclude that climatic conditions were warm and humid at higher elevations. These conditions were subject to slight thermal variations marked by alternations between thermophilous and steppe elements
(Fig. 6
). We also conclude that climatic conditions were warm and humid at higher elevations. These conditions were subject to slight thermal variations marked by alternations between thermophilous and steppe elements
(Fig. 6 ![]() ).
).
In addition to the P/C ratio, another indicator of this climatic evolution is provided by the Artemisia, Lygeum, Artemisia + Ephedra and S/T curves, which show an overall positive correlation with the δ18O isotope curve
(Fig. 6 ![]() ). The curve is considered the reference curve for global climate change, and oceanic isotope curves have long been correlated with cold/warm (glacial/interglacial) phases (Suballyova,
1997; Joannin,
2007; Popescu et al.,
2010; Suc et al., 2010,
2018b; Ohneiser et al.,
2015). The oxygen isotope ratio of seawater increases during cold phases. This increase in isotopic composition from the Early Pliocene to the Early Pleistocene and the expansion of Artemisia steppes are attributed to global cooling and the development of the first Northern Hemisphere ice caps at 2.6 Ma. The onset of these glaciations led to widespread xericity and cold conditions with highly contrasting seasons, while interglacials were milder and moister (Svenning,
2003; Suc et al., 2004, 2018b). These climatic trends were repeated during secondary fluctuations (Suc,
1978; Combourieu-Nebout et al.,
2000). During this time interval, the progressive aridification of the Mediterranean region also occurred (Suc,
1984; Leroy & Dupont,
1994; Fauquette et al., 1998b; Khélifi et al.,
2009; Popescu et al.,
2010), with the establishment of the Mediterranean-type seasonal precipitation rhythm (summer drought) at 3.4 Ma (Suc,
1984; Suc & Popescu,
2005; Barrón et al.,
2010; Jiménez-Moreno et al.,
2010). Simultaneously, the East African monsoon exerted a specific influence that preserved moisture in the northeastern Mediterranean region (Suc & Popescu,
2005; Popescu et al., 2006; Biltekin et al.,
2015).
). The curve is considered the reference curve for global climate change, and oceanic isotope curves have long been correlated with cold/warm (glacial/interglacial) phases (Suballyova,
1997; Joannin,
2007; Popescu et al.,
2010; Suc et al., 2010,
2018b; Ohneiser et al.,
2015). The oxygen isotope ratio of seawater increases during cold phases. This increase in isotopic composition from the Early Pliocene to the Early Pleistocene and the expansion of Artemisia steppes are attributed to global cooling and the development of the first Northern Hemisphere ice caps at 2.6 Ma. The onset of these glaciations led to widespread xericity and cold conditions with highly contrasting seasons, while interglacials were milder and moister (Svenning,
2003; Suc et al., 2004, 2018b). These climatic trends were repeated during secondary fluctuations (Suc,
1978; Combourieu-Nebout et al.,
2000). During this time interval, the progressive aridification of the Mediterranean region also occurred (Suc,
1984; Leroy & Dupont,
1994; Fauquette et al., 1998b; Khélifi et al.,
2009; Popescu et al.,
2010), with the establishment of the Mediterranean-type seasonal precipitation rhythm (summer drought) at 3.4 Ma (Suc,
1984; Suc & Popescu,
2005; Barrón et al.,
2010; Jiménez-Moreno et al.,
2010). Simultaneously, the East African monsoon exerted a specific influence that preserved moisture in the northeastern Mediterranean region (Suc & Popescu,
2005; Popescu et al., 2006; Biltekin et al.,
2015).
On the other hand, Suc (1984) and Popescu et al. (2010) considered the 3.37-2.6 Ma time interval as a transitional phase characterized by the emergence of a Mediterranean-type seasonal rhythm with summer droughts and cooler winters.
5.3.2. Sea-level variation
The distality markers (dinocysts, D/S ratio, Pinus and indeterminable Pinaceae, and halophytes) indicate that the depositional environment was epicontinental and strongly influenced by continental inputs. The inverse evolution of these distality indicators depicts a transition from proximal to distal neritic settings
(Fig. 6 ![]() ). These findings are fully consistent with those of earlier studies by Benzakour
(1992), Londeix et al.
(1999), and Feddi et al.
(2011). According to these authors, the distality index reflects the alternation of proximal neritic and distal neritic environments, suggesting relative sea-level variations linked to eustatic fluctuations. They also noted that the distality index curve parallels the isotope curves from oceanic environments. Additionally, Hilgen et al.
(2009) demonstrated that Pliocene marine sedimentation in the central Mediterranean (Sicily) was strongly controlled by the orbital parameter variability.
). These findings are fully consistent with those of earlier studies by Benzakour
(1992), Londeix et al.
(1999), and Feddi et al.
(2011). According to these authors, the distality index reflects the alternation of proximal neritic and distal neritic environments, suggesting relative sea-level variations linked to eustatic fluctuations. They also noted that the distality index curve parallels the isotope curves from oceanic environments. Additionally, Hilgen et al.
(2009) demonstrated that Pliocene marine sedimentation in the central Mediterranean (Sicily) was strongly controlled by the orbital parameter variability.
The influence of astronomical forcing could explain the aforementioned repetitive changes in vegetation in the Gulf of Hammamet. The decrease in thermophilous elements, dinocysts and Pinus + indeterminable Pinaceae, along with the increase in steppe elements and halophytes, can be attributed to the influence of a cold phase
(Fig. 6 ![]() ). On one hand, this phase led to a relative drop in sea level that favored the colonization of the coastline by halophytes and steppe elements; on the other hand, it caused a decrease of thermophilous elements (including megatherms, mega-mesotherms, and
mesotherms) (Fig. 6
). On one hand, this phase led to a relative drop in sea level that favored the colonization of the coastline by halophytes and steppe elements; on the other hand, it caused a decrease of thermophilous elements (including megatherms, mega-mesotherms, and
mesotherms) (Fig. 6 ![]() ). Conversely, the warm phases caused the depositional setting to shift offshore, the halophytes and steppe elements to decrease, and the thermophiles to proliferate
(Fig. 6
). Conversely, the warm phases caused the depositional setting to shift offshore, the halophytes and steppe elements to decrease, and the thermophiles to proliferate
(Fig. 6 ![]() ). It is worth recalling here that the Late Pliocene climate was characterized by cyclic variability, which led to these vegetation changes (dominated by herbaceous plants) (Tzedakis,
2007; Popescu et al.,
2010; Feddi et al.,
2011), mainly due to precession (Kloosterboer-Hoeve et al.,
2006; Tzedakis,
2007; Sánchez Gońi et al.,
2008), obliquity (Joannin et al.,
2007, 2008, 2011; Popescu et al.,
2010; Suc et al., 2010, 2018b), and eccentricity.
). It is worth recalling here that the Late Pliocene climate was characterized by cyclic variability, which led to these vegetation changes (dominated by herbaceous plants) (Tzedakis,
2007; Popescu et al.,
2010; Feddi et al.,
2011), mainly due to precession (Kloosterboer-Hoeve et al.,
2006; Tzedakis,
2007; Sánchez Gońi et al.,
2008), obliquity (Joannin et al.,
2007, 2008, 2011; Popescu et al.,
2010; Suc et al., 2010, 2018b), and eccentricity.
The palynological analysis conducted on the uppermost Messinian to Gelasian marine sediments of the Jiriba-1 borehole- dated by Benzakour (1992) and Londeix et al. (1999) based on planktonic foraminifera, and calibrated here using the timescales of Lourens et al. (2004), Iaccarino et al. (2007), and Lirer et al. (2019) - provided, for the first time, vegetation and climate information for the Gulf of Hammamet during this time interval.
Pollen analysis of the sedimentary formations from the Jiriba-1 borehole revealed a floral diversity that includes elements currently inhabiting the intertropical, subtropical, and warm-temperate regions of America and Asia, as well as elements currently living in the Euro-Mediterranean region. High percentages correspond to taxa representative of subdesertic environments and, more generally, xeric open plant formations.
The vegetation and climate reconstruction was based on the East China vegetation and climate model (Wang, 1961), whose ecological and climatic affinities closely resemble those of the Pliocene Gulf of Hammamet. Low-altitude regions were characterized by open subdesertic formations. At mid-altitude, mixed forest plant formations developed, while a tree grouping was established in-between, consisting of elements whose extant representatives display a Mediterranean distribution.
The evolutionary trajectories of the three curves of Lygeum, Artemisia, and the ratio of steppe elements to thermophilous elements (S/T) exhibit a strong concordance with the major climatic phases established by Suc and Zagwijn (1983), Suc et al. (1995), Popescu (2001), and Suc et al. (2018a), thus enabling the identification of corresponding phases in the Gulf of Hammamet region, namely:
This study further proposes a correlation between the climatic phases identified for the first time in the Gulf of Hammamet and the southwestern Mediterranean and those previously established by Suc et al. (2018a).
The climatic evolution indicated by the two curves of Artemisia and the steppe elements (Artemisia and Ephedra) from the Jiriba-1 borehole exhibits parallelisms with the δą⁸O isotope curve.
The distality markers indicate, on the one hand, that the depositional environment of the Jiriba-1 borehole was epicontinental and influenced by significant fluvial inputs, and on the other hand, an alternation between proximal and distal neritic conditions.
The repetitive changes in plant composition between the steppe elements (Artemisia and Ephedra), thermophiles (megatherms, mega-mesotherms, and mesotherms), and halophytes in the Gulf of Hammamet could be explained by the impact of astronomical forcing.
This research benefited from the technical equipment of the Laboratory of Geosciences, Geomatics, and Environment, Geology Department, Faculty of Sciences Ben M'Sick, Hassan II University of Casablanca (Morocco). We express our sincere gratitude to Alberto Collareta for his diligent editorial oversight and invaluable suggestions throughout the peer-review process, and to Bruno Granier for his insightful and constructive comments. We thank the language editor for their careful review and the anonymous reviewers for their thoughtful feedback, which significantly improved the quality of this manuscript.
Abidi O., Inoubli M.H., Sebei K., Boussiga H., Amiri A., Hamdi I. & Ben Salem A.H. (2015).- Geodynamic framework and petroleum potential of the Cap Bon-Gulf of Hammamet Province-Tunisia. In: International Conference on Engineering Geophysics, Al Ain, United Arab Emirates, 15-18 November 2015.- SEG Global Meeting Abstracts, p. 264-266.
Achalhi M., Münch P., Cornée J.J., Azdimousa A., Melinte-Dobrinescu M., Quillévéré F., Drinia H., Fauquette S., Jiménez-Moreno G., Merzeraud G., Moussa A.B., El Karim Y. & Feddi N. (2016).- The late Miocene Mediterranean-Atlantic connections through the north Rifian corridor: New insights from the Boudinar and Arbaa Taourirt basins (northeastern Rif, Morocco).- Palćogeography, Palćoclimatology, Palćoecology, vol. 459, p. 131-152.
Argnani A. & Torelli L. (2001).- The Pelagian shelf and its graben system. In: Peri-Tethys Memoir 6: Peri-Tethyan rift/wrench basins and passive margins.- Mémoires du Muséum national d'Histoire naturelle, Paris, vol. 186, p. 529-544.
Bache F., Popescu S.-M., Rabineau M., Gorini C., Suc J.-P., Clauzon G., Olivet J.L., Rubino J.L., Melinte-Dobrinescu M.C., Estrada F., Londeix L., Armijo R., Meyer B., Jolivet L., Jouannic G., Leroux E., Aslanian D., Dos Reis A.T., Mocochain L., Dumurdžanov N., Zagorchev I., Lesić V., Tomić D., Namik Çağatay N., Brun J.-P., Sokoutis D., Csato I., Ucarkus G. & Çakir Z. (2012).- A two-step process for the reflooding of the Mediterranean after the Messinian Salinity Crisis.- Basin Research, vol. 24, no 2, p. 125-153.
Bachiri Taoufiq N. (2000, unpublished).- Les environnements marins et continentaux du corridor rifain au Miocčne supérieur d'aprčs la palynologie.- Thčse d'État es Sciences, Université Hassan II, Casablanca, 206 p.
Bachiri Taoufiq N., Barhoun N. & Suc J.-P. (2008).- Les environnements continentaux du corridor rifain (Maroc) au Miocčne supérieur d'aprčs la palynologie.- Geodiversitas, Paris, vol. 30, no 1, p. 41-58.
Bahaj H., Barhoun N., Bachiri Taoufiq N., Rahmouna J., Targhi S., Berry N., Suc J.-P. & Popescu S.-M. (2024).- Paleoenvironmental changes preceding the onset of the Messinian salinity crisis in the western Mediterranean Sea (pre-evaporitic Messinian of the Melilla-Nador Basin, NE Morocco).- Carnets Geol., Madrid, vol. 24, no. 1, p. 1-27. DOI: 10.2110/carnets.2024.2401
Barhoun N. & Bachiri Taoufiq N. (2008).- Événements biostratigraphiques et environnementaux enregistrés dans le corridor sud rifain (Maroc septentrional) au Miocčne supérieur avant la crise de salinité messinienne.- Geodiversitas, vol. 30, no. 1, p. 21-40.
Barrón E., Rivas-Carballo R., Postigo-Mijarra J., Alcalde-Olivares C., Vieira M., Castro L., Pais J. & Valle-Hernandez M. (2010).- The Cenozoic vegetation of the Iberian Peninsula: A synthesis.- Review of Palaeobotany and Palynology, vol. 162, no. 3, p. 382-402.
Bartoli G., Sarnthein M., Weinelt M., Erlenkeuser H., Garbe-Schönberg D. & Lea D. W. (2005).- Final closure of Panama and the onset of northern hemisphere glaciation.- Earth and Planetary Science Letters, vol. 237, no. 1-2, p. 33-44.
Bédir M. (1995).- Mécanismes géodynamiques des bassins associés aux couloirs de coulissement de la marge atlasique de la Tunisie. Séismo-tectonique et implications pétroličres.- Thčse de Doctorat, Université de Tunis El Manar, 344 p. URL: https://hal.science/tel-01862051
Bédir M., Tlig S., Bobier C. & Issaoui N. (1996).- Sequence stratigraphy, Basin dynamics, and petroleum geology of the Miocene from eastern Tunisia.- AAPG Bulletin, Boulder - CO, vol. 80, no. 1, p. 63-80.
Bédir M., Zargouni F., Tlig S. & Bobier C. (1992).- Subsurface geodynamics and petroleum geology of transform margin basins in the Sahel of Mahdia and El Jem (eastern Tunisia).- AAPG Bulletin, Boulder - CO, vol. 76, no. 9, p. 1417-1442.
Ben Ayed N., Viguier C. & Bobier C. (1983).- Les éléments structuraux récents essentiels de la Tunisie nord-orientale.- Notes du Service géologique, Tunis, no. 47, p. 5-19.
Ben Brahim G., Brahim N. & Turki F. (2013).- Neogene tectonic evolution of the Gulf of Hammamet area, Northeast Tunisia offshore.- Journal of African Earth Sciences, vol. 81, p. 16-27.
Ben Moktar N. & Mannaď-Tayech B. (2012).- Reconstitution de la végétation et du climat durant le Miocčne dans le bassin de Saouaf (Tunisie centro-nord-orientale).- Geodiversitas, Paris, vol. 34, no. 2, p. 445-456.
Bensalem H. (1992, unpublished).- Contribution ŕ la connaissance de la géologie du Cap Bon : Stratigraphie, tectonique et sédimentologie.- Thčse de Spécialité, Université de Tunis, 155 p.
Benzakour M. (1992).- Palynologie du Néogčne dans la région du détroit de Sicile, Méditerranée centrale.- Thčse de Doctorat en Sciences de l'Environnement, Université du Québec, Montréal, 176 p.
Berg B.C.J. van den (2016).- Late Miocene astrochronology and basin evolution of the Atlantic side of the Betic Corridor linked to the Messinian salinity crisis.- Tesis doctoral, Universidad de Salamanca, 174 p. URL: https://hdl.handle.net/10366/132993
Berger A. & Morel P. (1992).- Le climat de la Terre : Un passé pour quel avenir?- De Boeck Wesmael, Bruxelles, 479 p.
Berggren W.A., Kent D.V., Flynn J.J. & Van Couvering J.A. (1985).- Cenozoic geochronology.- Geological Society of America Bulletin, Boulder - CO, vol. 96, no. 11, p. 1407-1418.
Bernatchez P., Fraser C., Friesinger S., Jolivet Y., Dugas S., Drejza S. & Morissette A. (2008).- Sensibilité des côtes et vulnérabilité des communautés du golfe du Saint-Laurent aux impacts des changements climatiques.- Ouranos, Montréal, 48 p. URL: https://www.bibliotheque.assnat.qc.ca/DepotNumerique_v2/AffichageFichier.aspx?idf=141262
Berry N., Targhi S., Barhoun N., Bachiri Taoufiq N., Essamoud R., Flecker R., Ducassou E., Williams T. & The IODP Expedition 401 Science Party (2025).- Reconstruction of environmental conditions in the Alboran Sea during the Zanclean: Insights from a preliminary palynological analysis.- 57th Annual Meeting AASP - The Palynological Society, Rabat, Morocco (22-26 April 2025), p. 142-143.
Bertini A. (1994).- Palynological investigations on Upper Neogene and Lower Pleistocene sections in Central and Northern Italy.- Memorie della Societa Geologica Italiana, Roma, vol. 48, p. 431-443.
Bertini A. (2001).- Pliocene climatic cycles and altitudinal forest development from 2.7 Ma in the Northern Apennines (Italy): Evidence from the pollen record of the Stirone section (~5.1 to ~2.2 Ma).- Geobios, Villeurbanne, vol. 34, no. 3, p. 253-265.
Bertini A. (2010).- Pliocene to Pleistocene palynoflora and vegetation in Italy: State of the art.- Quaternary International, vol. 225, no. 1, p. 5-24.
Bertini A. (2013).- Climate and vegetation in the Upper Valdarno Basin (central Italy) as a response to Northern Hemisphere insolation forcing and regional tectonics in the late Pliocene-early Pleistocene.- Italian Journal of Geosciences, Roma, vol. 132, no. 1, p. 137-148.
Bertini A. & Combourieu-Nebout N. (2023).- Piacenzian to late Pleistocene flora and vegetation in Italy: A moving sketch.- Alpine and Mediterranean Quaternary, Parma, vol. 36, no. 1, p. 91-119.
Bertini A., Marino M., Combourieu-Nebout N., Girone A., Maiorano P., Niccolini G. & Suc J.-P. (2025).- Unravelling land-sea correlations during the Pliocene-Pleistocene in the Mediterranean area.- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 64, p. 289-312.
Besčme P. & Kamoun Y. (1988).- Le Messinien marin de Ksour Essaf (Sahel, Tunisie orientale), une étude stratigraphique, sédimentologique et paléoécologique.- Revue des Sciences de la Terre, Tunis, vol. 8, p. 129-142.
Besčme P. & Lajmi T. (1981).- Le Pliocčne marin du lac Ichkeul (coupe du Jebel Merzaguine) : Son contexte géologique, son évolution sédimentologique, son contenu microfaunistique.- Actes du Premier Congrčs National des Sciences de la Terre, Tunis, p. 69-83.
Bessedik M. (1985).- Reconstitution des environnements miocčnes des régions nord-ouest méditerranéennes ŕ partir de la palynologie.- Geobios, Villeurbanne, vol. 18, no. 6, p. 905-905.
Biltekin D., Popescu S.-M., Suc J.-P., Quézel P., Jiménez Moreno G., Yavuz N. & Çagatay M.N. (2015).- Anatolia: A long-time plant refuge area documented by pollen records over the last 23 million years.- Review of Palaeobotany and Palynology, vol. 215, p. 1-22.
Bismuth H. & Benzarti R. (1982, unpublished).- Étude biostratigraphique du sondage Jiriba 1 (JRI.1) (Golfe de Hammamat).- SEREPT, Rapport Interne (Tunis), GN2470 (1982-Ex.11-09), 13 p.
Bizon G., Bizon J., Burollet P.F. & Jelisejeff A. (1980).- Le Pliocčne inférieur de Raf-Raf (Tunisie Nord-Orientale). Comparaison avec les séries de męme âge observées en Espagne méridionale et dans les îles ioniennes.- Annales des Mines et de la Géologie, Tunis, vol. 3, no. 28, p. 301-313.
Blanc P.L. (2002).- The opening of the Plio-Quaternary Gibraltar Strait: Assessing the size of a cataclysm.- Geodinamica Acta, vol. 15, no. 5-6, p. 303-317.
Blanc P.L., Fontugne M.R. & Duplessy J.C. (1983).- The time-transgressive initiation of boreal ice-caps: Continental and oceanic evidence reconciled.- Palćogeography, Palćoclimatology, Palćoecology, vol. 42, no. 3-4, p. 211-224.
Boccaletti M., Cello G. & Tortorici L. (1987).- Transtensional tectonics in the Sicily Channel.- Journal of Structural Geology, vol. 9, no. 7, p. 869-876.
Bouaziz S., Barrier E., Soussi M., Turki M.M. & Zouari H. (2002).- Tectonic evolution of the northern African margin in Tunisia from paleostress data and sedimentary record.- Tectonophysics, vol. 357, no. 1-4, p. 227-253.
Burollet P.F. (1991).- Structures and tectonics of Tunisia.- Tectonophysics, vol. 195, no. 2-4, p. 359-369.
Capella W., Barhoun N., Flecker R., Hilgen F.J., Kouwenhoven T., Matenco L.C., Sierro F.J., Tulbure M.A., Yousfi M.Z. & Krijgsman W. (2018).- Palaeogeographic evolution of the late Miocene Rifian Corridor (Morocco): Reconstructions from surface and subsurface data.- Earth-Science Reviews, vol. 180, p. 37-59.
Capella W., Flecker R., Hernández-Molina F.J., Simon D., Meijer P.T., Rogerson M., Sierro F.J. & Krijgsman W. (2019).- Mediterranean isolation preconditioning the Earth System for late Miocene climate cooling.- Scientific Reports, vol. 9, no. 1, 8 p. DOI: 10.1038/s41598-019-40208-2
Catalano R., Di Stefano P., Sulli A. & Vitale F.P. (1996).- Paleogeography and structure of the central Mediterranean; Sicily and its offshore area.- Tectonophysics, vol. 260, no. 4, p. 291-323.
Chihi L. & Philip H. (1999).- Le bloc atlasopélagien : Place et évolution géodynamique dans le contexte subduction-collision de la Méditerranée centrale (Afrique du Nord-Sicile) du Miocčne au Quaternaire.- Notes du Service géologique de Tunisie, Tunis, vol. 65, p. 49-61.
Chikhi H. (1992a).- Une palynoflore méditerranéenne ŕ subtropicale au Messinien pré-évaporitique en Algérie.- Géologie Méditerranéenne, Marseille, t. XIX, no. 1, p. 19-30. URL: https://www.persee.fr/doc/geolm_0397-2844_1992_num_19_1_1466
Chikhi H. (1992b).- Palynoflore du Messinien infra-évaporitique de la série marno-diatomitique de Sahaouria (Beni-Chougrane) et de Chabet Bou Seter (Tessala), bassin du Chelif, Algérie.- Thčse de Magister, Université d'Oran, 164 p. (Pls. XII-XIII). URL: https://dspace.univ-tlemcen.dz/handle/112/6772
Clauzon G., Suc J.-P., Do Couto D., Jouannic G., Melinte-Dobrinescu M. C., Jolivet L., Lebret N., Mocochain L., Popescu S.-M., Martinell J., Domenech R., Rubino J.-L., Gumiaux C., Warny S., Gorini C., Bache F., Rabineau M. & Estrada F. (2015).- New insights on the Sorbas Basin (SE Spain): The onshore reference of the Messinian Salinity Crisis.- Marine and Petroleum Geology, vol. 66, p. 71-100.
Combourieu-Nebout N., Bertini A., Russo-Ermolli E., Peyron O., Klotz S., Montade V., Fauquette S., Allen J., Fusco F., Goring S., Huntley B., Joannin S., Lebreton V., Magri D., Martinetto E., Orain R. & Sadori L. (2015).- Climate changes in the central Mediterranean and Italian vegetation dynamics since the Pliocene.- Review of Palaeobotany and Palynology, vol. 218, p. 127-147.
Combourieu-Nebout N., Fauquette S. & Quézel P. (2000).- What was the late Pliocene Mediterranean climate like? A preliminary quantification from vegetation.- Bulletin de la Société géologique de France, Paris, vol. 171, no. 2, p. 271-277.
Cornée J. J., Münch P., Achalhi M., Merzeraud G., Azdimousa A., Quillévéré F., Melinte-dobrinescu M., Chaix C., Ben Moussa A., Lofi J., Séranne M. & Moissette P. (2016).- The Messinian erosional surface and early Pliocene reflooding in the Alboran Sea: New insights from the Boudinar basin, Morocco.- Sedimentary Geology, vol. 333, p. 115-129.
Cour P. & Duzer D. (1978).- La signification climatique, édaphique et sédimentologique des rapports entre taxons en analyse pollinique.- Annales des Mines de Belgique, Bruxelles, no. 7-8, p. 825-834.
De Schepper S., Head M.J. & Groeneveld J. (2009).- North Atlantic Current variability through marine isotope stage M2 (circa 3.3 Ma) during the mid-Pliocene.- Paleoceanography, Washington - DC, vol. 24, no. 4, 17 p. DOI: 10.1029/2008PA001725
De Schepper S., Groeneveld J., Naafs B.D.A., Van Renterghem C., Hennissen J., Head M.J., Louwye S. & Fabian K. (2013).- Northern Hemisphere Glaciation during the Globally Warm Early Late Pliocene.- PLOS One, Washington - DC, vol. 8, no. 12, article e81508, 15 p. DOI: 10.1371/journal.pone.0081508
Derbel-Damak F. & Zaghbib-Turki D. (2002).- Identification des zones biostratigraphiques méditerranéennes dans le Pliocčne du Cap-Bon (Tunisie).- Geobios, vol. 35, no. 2, p. 253-264.
Dijk G. van, Maars J., Andreetto F., Hernández-Molina F.J., Rodríguez-Tovar F.J. & Krijgsman W. (2023).- A terminal Messinian flooding of the Mediterranean evidenced by contouritic deposits on Sicily.- Sedimentology, vol. 70, no. 4, p. 1195-1223.
Diniz F. (1984).- Apports de la palynologie ŕ la connaissance du Pliocčne portugais : Rio Maior : Un bassin de référence pour l'histoire de la flore, de la végétation et du climat de la façade atlantique de l'Europe méridionale.- Thčse d'État, Université Montpellier II, 230 p.
Dlala M. (1994).- Evidence for interchange of the stress (σ1; σ2) and its geodynamic implication.- Africa Geoscience Review, Paris, vol. 1, p. 533-545.
Dlala M. (1995).- Évolution géodynamique et tectoniques superposées en Tunisie : Sur l'implication tectonique récente et la sismicité.- Thčse d'État, Université de Tunis El Manar, 389 p.
Do Couto D. (2014).- Évolution géodynamique de la Mer d'Alboran par l'étude des bassins sédimentaires.- Thčse de Doctorat, Université Pierre et Marie Curie-Paris VI, 502 p. URL: https://hal-bioemco.ccsd.cnrs.fr/THESES-UPMC/tel-01148777
Drivaliari A., Ticleanu N., Marinescu F., Marunteanu M. & Suc J.-P. (1999).- A Pliocene climatic record at Ticleni (southwestern Romania). In: Wrenn J.H., Suc J.-P. & Leroy S.A.G. (eds.), The Pliocene: Time of change.- American Association of Stratigraphic Palynologists Foundation, Dallas - TX, p. 103-108.
Estrada F., Ercilla G., Gorini C., Alonso B., Vázquez J.T., García-Castellanos D., Juan C., Maldonado A., Ammar A. & Elabbassi M. (2011).- Impact of pulsed Atlantic water inflow into the Alboran Basin at the time of the Zanclean flooding.- Geo-Marine Letters, vol. 31, no. 5, p. 361-376.
Faccenna C., Piromallo C., Crespo-Blanc A., Jolivet L. & Rossetti F. (2004).- Lateral slab deformation and the origin of the western Mediterranean arcs.- Tectonics, vol. 23, no. 1, article TC1012, 21 p. DOI: 10.1029/2002TC001488
Fauquette S., Guiot J. & Suc J.-P. (1998a).- A method for climatic reconstruction of the Mediterranean Pliocene using pollen data.- Palćogeography, Palćoclimatology, Palćoecology, vol. 144, no. 1-2, p. 183-201.
Fauquette S., Quézel P., Guiot J. & Suc J.-P. (1998b).- Signification bioclimatique de taxons-guides du Pliocčne méditerranéen.- Geobios, Villeurbanne, vol. 31, no. 2, p. 151-169.
Fauquette S., Suc J.-P., Guiot J., Diniz F., Feddi N., Zheng Z., Bessais E. & Drivaliari A. (1999).- Climate and biome in the West Mediterranean area during the Pliocene.- Palćogeography, Palćoclimatology, Palćoecology, vol. 152, no. 1-2, p. 15-36.
Fauquette S., Suc J.-P., Jiménez-Moreno G., Micheels A., Jost A., Favre E., Bachiri Taoufiq N., Bertini A., Clet-Pellerin M., Diniz F., Farjanel G., Feddi N. & Zheng Z. (2007).- Latitudinal climatic gradients in western European and Mediterranean regions from the Mid-Miocene (c. 15 Ma) to the Mid-Pliocene (c. 3.5 Ma) as quantified from pollen data. In: Williams M., Haywood A., Gregory J. & Schmidt D. (eds.), Deep time perspectives on climate change: Marrying the signal from computer models and biological proxies.- The Micropaleontological Society, Special Publications, London, p. 481-502.
Feddi N., Fauquette S. & Suc J.-P. (2011).- Histoire plio-pléistocčne des écosystčmes végétaux de Méditerranée sud-occidentale : Apport de l'analyse pollinique de deux sondages en mer d'Alboran.- Geobios, Villeurbanne, vol. 44, no. 1, p. 57-69.
Flecker R., Krijgsman W., Capella W., Castro Martins C. de, Demitrieva E., Mayser J.-P., Marzocchi A., Modestu S., Ochoa Lozano D., Simon D., Tulbure M., Berg B. van den, Schee M. van der, Lange G. de, Ellam R., Govers R., Gutjahr M., Hilgen F., Kouwenhoven T., Lofi J., Meijer P., Sierro F.J., Bachiri Taoufiq N., Barhoun N., Chakor Alami A., Chacon B., Flores J.A., Gregory J., Howard J., Lunt D., Ochoa M., Pancost R., Vincent S. & Yousfi M.Z. (2015).- Evolution of the Late Miocene Mediterranean Atlantic gateways and their impact on regional and global environmental change.- Earth-Science Reviews, vol. 150, p. 365-392.
Frigui M., Youssef M.B. & Ouaja M. (2016).- Evidences of "Lago-Mare" episode around the Messinian-Pliocene boundary in eastern Tunisia (central Mediterranean).- Journal of African Earth Sciences, vol. 123, p. 57-74.
Gaaloul N. & Razgallah S. (2008).- Les paléoenvironnements du Messinien de la Tunisie orientale (Sahel et Golfe de Hammamet).- Geo-Eco-Trop, Ličge, vol. 32, p. 91-100.
García-Castellanos D., Estrada F., Jiménez-Munt I., Gorini C., Fernández M., Verges J. & De Vicente R. (2009).- Catastrophic flood of the Mediterranean after the Messinian salinity crisis.- Nature, vol. 462, no. 7274, p. 778-781.
Gardiner W., Grasso M. & Sedgeley D. (1995).- Plio-Pleistocene fault movement as evidence for mega-block kinematics within the Hyblean-Malta plateau, Central Mediterranean.- Journal of Geodynamics, vol. 19, no. 1, p. 35-51.
Haller P. (1983).- Structure profonde du Sahel tunisien : Interprétation géodynamique.- Thčse d'État, Université de Franche-Comté, Besançon, 163 p.
Haug G.H. & Tiedemann R. (1998).- Effect of the formation of the Isthmus of Panama on Atlantic Ocean thermohaline circulation.- Nature, vol. 393, no. 6686, p. 673-676.
Heusser L. & Balsam W.L. (1977).- Pollen distribution in the northeast Pacific Ocean.- Quaternary Research, Cambridge (UK), vol. 7, no. 1, p. 45-62.
Hilgen F.J. (1990).- Closing the gap in the Plio-Pleistocene boundary stratotype sequence of Crotone (southern Italy).- Newsletters on Stratigraphy, vol. 32, p. 43-51.
Hilgen F.J. (1991).- Extension of the astronomically calibrated (polarity) time scale to the Miocene/Pliocene boundary.- Earth and Planetary Science Letters, vol. 107, no. 2, p. 349-368.
Hilgen F.J., Abels H.A., Iaccarino S., Krijgsman W., Raffi I., Sprovieri R., Turco E. & Zachariasse W.J. (2009).- The global stratotype section and point (GSSP) of the Serravallian Stage (Middle Miocene).- Episodes, Seoul, vol. 32, no. 3, p. 152-166.
Hsü K.J., Ryan W.B. & Cita M.B. (1973).- Late Miocene desiccation of the Mediterranean.- Nature, vol. 242, no. 5395, p. 240-244.
Iaccarino S.M., Premoli Silva I., Biolzi M., Foresi L.M., Lirer F., Turco E. & Petrizzo M.R. (2007).- Practical manual of Neogene planktonic foraminifera. In: International school on planktonic Foraminifera (Neogene planktonic Foraminifera).- Universitŕ di Perugia Press, 180 p.
Jiménez-Moreno G., Alçiçek H., Alçiçek M.C., Hoek Ostende L. van den & Wesselingh F.P. (2015).- Vegetation and climate changes during the late Pliocene and early Pleistocene in SW Anatolia, Turkey.- Quaternary Research, vol. 84, no. 3, p. 448-456.
Jiménez-Moreno G., Fauquette S. & Suc J.-P. (2008).- Vegetation, climate, and palaeoaltitude reconstructions of the Eastern Alps during the Miocene based on pollen records from Austria, Central Europe.- Journal of Biogeography, vol. 35, no. 9, p. 1638-1649.
Jiménez-Moreno G., Fauquette S. & Suc J.-P. (2010).- Miocene to Pliocene vegetation reconstruction and climate estimates in the Iberian Peninsula from pollen data.- Review of Palaeobotany and Palynology, vol. 162, no. 3, p. 403-415.
Jiménez-Moreno G., Leeuw A. de, Mandic O., Harzhauser M., Pavelić D., Krijgsman W. & Vranjković A. (2009).- Integrated stratigraphy of the Early Miocene lacustrine deposits of Pag island (SW Croatia): Palaeovegetation and environmental changes in the Dinaride Lake System.- Palćogeography, Palćoclimatology, Palćoecology, vol. 280, no. 1-2, p. 193-206.
Jiménez-Moreno G., Pérez-Asensio J.N., Larrasoańa J.C., Aguirre J., Civis J., Rivas-Carballo M.R., Valle-Hernández M.F. & González-Delgado J.A. (2013).- Vegetation, sea-level, and climate changes during the Messinian salinity crisis.- GSA Bulletin, Boulder - CO, vol. 125, no. 3-4, p. 432-444.
Joannin S. (2007).- Changements climatiques en Méditerranée ŕ la transition Pléistocčne inférieur-moyen : Pollens, isotopes stables et cyclostratigraphie.- Thčse, Diplôme de Doctorat, Université Claude Bernard-Lyon I, 220 p. URL: https://hal.science/tel-00347549/
Joannin S., Bassinot F., Nebout N. C., Peyron O. & Beaudouin C. (2011).- Vegetation response to obliquity and precession forcing during the Mid-Pleistocene Transition in Western Mediterranean region (ODP site 976).- Quaternary Science Reviews, vol. 30, no. 3-4, p. 280-297.
Joannin S., Ciaranfi N. & Stefanelli S. (2008).- Vegetation changes during the late Early Pleistocene at Montalbano Jonico (Province of Matera, southern Italy) based on pollen analysis.- Palćogeography, Palćoclimatology, Palćoecology, vol. 270, no. 1-2, p. 92-101.
Joannin S., Cornée J.-J., Moissette P., Suc J.-P., Koskeridou E., Lécuyer C., Buisine C., Kouli K. & Ferry S. (2007).- Changes in vegetation and marine environments in the eastern Mediterranean (Rhodes, Greece) during the Early and Middle Pleistocene.- Journal of the Geological Society, London, vol. 164, no. 6, p. 1119-1131.
Jolivet L., Augier R., Robin C., Suc J.-P. & Rouchy J.-M. (2006).- Lithospheric-scale geodynamic context of the Messinian salinity crisis.- Sedimentary Geology, vol. 188-189, p. 9-33.
Jolivet L. & Faccenna C. (2000).- Mediterranean extension and the Africa-Eurasia collision.- Tectonics, vol. 19, no. 6, p. 1095-1106. DOI: 10.1029/2000TC900018
Kaboth-Bahr S., Bahr A., Stepanek C., Catunda M.C.A., Karas C., Ziegler M., García-Gallardo A. & Grunert P. (2021).- Mediterranean heat injection to the North Atlantic delayed the intensification of Northern Hemisphere glaciations.- Communications Earth & Environment, vol. 2, no. 1, article 158, 9 p. DOI: 10.1038/s43247-021-00232-5
Kacem J. (2004).- Étude sismotectonique et évaluation de l'aléa sismique régional du Nord-Est de la Tunisie : Apport de la sismique réflexion dans l'identification des sources sismogéniques.- Thčse de Doctorat d'Université, Université de Tunis El Manar, 268 p.
Karas C., Khélifi N., Bahr A., Naafs B.D.A., Nürnberg D. & Herrle J.O. (2020).- Did North Atlantic cooling and freshening from 3.65-3.5 Ma precondition Northern Hemisphere ice sheet growth?.- Global and Planetary Change, vol. 185, article 103085, 7 p.
Khélifi N., Sarnthein M., Andersen N., Blanz T., Frank M., Garbe-Schönberg D., Haley B.A., Stumpf R. & Weinelt M. (2009).- A major and long-term Pliocene intensification of the Mediterranean outflow, 3.5-3.3 Ma ago.- Geology, Boulder - CO, vol. 37, no. 9, p. 811-814.
Kloosterboer-Hoeve (van) M.L., Steenbrink J., Visscher H. & Brinkhuis H. (2006).- Millennial-scale climatic cycles in the Early Pliocene pollen record of Ptolemais, northern Greece.- Palćogeography, Palćoclimatology, Palćoecology, vol. 229, no. 4, p. 321-334.
Krijgsman W., Capella W., Simon D., Hilgen F.J., Kouwenhoven T.J., Meijer P.T., Sierro F.J., Tulbure M.A., Berg B.C.J. van den, Schee M. van der & Flecker R. (2018).- The Gibraltar corridor: Watergate of the Messinian salinity crisis.- Marine Geology, vol. 403, p. 238-246.
Krijgsman W., Hilgen F.J., Raffi I., Sierro F.J. & Wilson D.S. (1999).- Chronology, causes and progression of the Messinian salinity crisis.- Nature, vol. 400, article 6745, p. 652-655.
Krijgsman W. & Meijer P.T. (2008).- Depositional environments of the Mediterranean "Lower Evaporites" of the Messinian salinity crisis: Constraints from quantitative analyses.- Marine Geology, vol. 253, no. 3-4, p. 73-81.
Laan E. van der, Snel E., Kaenel E. de, Hilgen F.J. & Krijgsman W. (2006).- No major deglaciation across the Miocene-Pliocene boundary: Integrated stratigraphy and astronomical tuning of the Loulja sections (Bou Regreg area, NW Morocco).- Paleoceanography, vol. 21, no. 3, article PA3011, 27 p. DOI: 10.1029/2005PA001193
Laffitte R. & Dumon É. (1948).- Plissement Pliocčne supérieur et mouvement quaternaire en Tunisie.- Comptes Rendus de l'Académie des Sciences, Paris, t. 227, p. 138-140. 188.
Langereis C.G. & Hilgen F.J. (1991).- The Rossello composite: A Mediterranean and global reference section for the Early to early Late Pliocene.- Earth and Planetary Science Letters, vol. 104, no. 2-4, p. 211-225.
Leroy S. & Dupont L. (1994).- Development of vegetation and continental aridity in northwestern Africa during the Late Pliocene: The pollen record of ODP Site 658.- Palćogeography, Palćoclimatology, Palćoecology, vol. 109, no. 2-4, p. 295-316.
Lirer F., Foresi L.M., Iaccarino S.M., Salvatorini G., Turco E., Cosentino C., Sierro F.J. & Caruso A. (2019).- Mediterranean Neogene planktonic foraminifer biozonation and biochronology.- Earth-Science Reviews, vol. 196, article 102869, 36 p.
Lisiecki L.E. & Raymo M.E. (2005).- A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records.- Paleoceanography, vol. 20, no. 1, 17 p. DOI: 10.1029/2004PA001071
Lisiecki L.E. & Raymo M.E. (2007).- Plio-Pleistocene climate evolution: Trends and transitions in glacial cycle dynamics.- Quaternary Science Reviews, vol. 26, no. 1-2, p. 56-69.
Loget N. & Driessche J. van den (2006).- On the origin of the Strait of Gibraltar.- Sedimentary Geology, vol. 188, p. 341-356.
Londeix L., Benzakour M., De Vernal A., Turon J. L. & Suc J.-P. (1999).- Late Neogene dinoflagellate cyst assemblages from the Strait of Sicily, Central Mediterranean Sea: Paleoecological and biostratigraphical implications. In: Wrenn J.H., Suc J.-P. & Leroy S.A.G. (eds.), The Pliocene: Time of change.- American Association of Stratigraphic Palynologists Foundation, Dallas - TX, p. 65-91.
Lourens L., Hilgen F.J., Laskar J., Shackleton N.J. & Wilson D. (2004).- The Neogene period. In: Gradstein F., Ogg J. & Smith A. (eds.), A Geologic Time Scale.- Cambridge University Press, p. 409-440.
Mannaď-Tayech B. (2006).- Les séries silicoclastiques miocčnes du Nord-Est au Sud-Ouest de la Tunisie : Une mise au point.- Geobios, Villeurbanne, vol. 39, no. 1, p. 71-84.
Martín Martín J.M., Puga Bernabéu Á., Aguirre Rodríguez J. & Braga Alarcón J. (2014).- Miocene Atlantic-Mediterranean seaways in the Betic Cordillera (southern Spain).- Revista de la Sociedad Geológica de Espańa, Madrid, vol. 27, no. 1, p. 175-186.
McClymont E.L., Ho S.L., Ford H.L., Bailey I., Berke M.A., Bolton C.T., De Schepper S., Grant G.R., Groeneveld J., Inglis G.N., Karas C., Patterson M.O., Swann G.E.A., Thirumalai K., White S.M., Alonso-Garcia M., Anand P., Hoogakker B.A.A., Littler K., Petrick B.F., Risebrobakken B., Abell J.T., Crocker A.J., De Graaf F., Feakins S.J., Hargreaves J.C., Jones C.L., Markowska M., Ratnayake A.S., Stepanek C. & Tangunan D. (2023).- Climate evolution through the onset and intensification of Northern Hemisphere Glaciation.- Reviews of Geophysics, vol. 61, no. 3, article e2022RG000793, 43 p. DOI: 10.1029/2022RG000793
Mehta A.V. & Yang S. (2008).- Precipitation climatology over Mediterranean Basin from ten years of TRMM measurements.- Advances in Geosciences, vol. 17, p. 87-91. URL: 10.5194/adgeo-17-87-2008
Mejri L. (2012).- Tectonique Quaternaire, paléosismicité et sources sismogéniques en Tunisie Nord-orientale : Étude de la faille d'Utique.- Thčse de Doctorat de l'Université de Toulouse, Université Toulouse 3 Paul Sabatier, 174 p. URL: https://utheme.univ-tlse3.fr/s/fr/item/1669
Moissette P., Cornée J. J., Mannaď-Tayech B., Rabhi M., André J.-P., Koskeridou E. & Méon H. (2010).- The western edge of the Mediterranean Pelagian Platform: A Messinian mixed siliciclastic-carbonate ramp in northern Tunisia.- Palćogeography, Palćoclimatology, Palćoecology, vol. 285, no. 1-2, p. 85-103.
Mudelsee M. & Raymo M.E. (2005).- Slow dynamics of the Northern Hemisphere glaciation.- Paleoceanography, vol. 20, no. 4, 14 p. DOI: 10.1029/2005PA001153
Nebout N.C. & Grazzini C. V. (1991).- Late Pliocene northern hemisphere glaciations: The continental and marine responses in the central Mediterranean.- Quaternary Science Reviews, vol. 10, no. 4, p. 319-334.
Nix H.A. (1982).- Environmental determinants of biogeography and evolution in Terra Australis. In: Barker W.R. & Greenslade P.J.M. (eds.), Evolution of the flora and fauna of arid Australia.- Peacock Publications, Adelaide, p. 47-66.
Ohneiser C., Florindo F., Stocchi P., Roberts A.P., DeConto R.M. & Pollard D. (2015).- Antarctic glacio-eustatic contributions to late Miocene Mediterranean desiccation and reflooding.- Nature Communications, vol. 6, no. 1, article 8765, 10 p.
Patriat M., Ellouze N., Dey Z., Gaulier J.M. & Ben Kilani H. (2003).- The Hammamet, Gabes and Chotts (Tunisia): A review of the subsidence history.- Sedimentary Geology, vol. 156, no. 1-4, p. 241-262.
Piquet A., Tricart P., Guraud R., Laville E., Bouaziz S., Amrhar M. & Ait R. (2002).- The Mesozoic-Cenozoic Atlas belt (North Africa): An overview.- Geodinamica Acta, vol. 15, no. 3, p. 185-208.
Popescu S.-M. (2001).- Végétation, climat et cyclostratigraphie en Paratéthys centrale au Miocčne supérieur et au Pliocčne inférieur d'aprčs la palynologie.- Thčse, Diplôme de Doctorat, Université Claude Bernard-Lyon I, 233 p. URL: https://theses.hal.science/tel-00350111
Popescu S.-M. (2006).- Late Miocene and early Pliocene environments in the southwestern Black Sea region from high-resolution palynology of DSDP Site 380A (Leg 42B).- Palćogeography, Palćoclimatology, Palćoecology, vol. 238, no. 1-4, p. 64-77.
Popescu S.-M., Biltekin D., Winter H., Suc J.-P., Melinte-Dobrinescu M.C., Klotz S., Combourieu-Nebout N., Rabineau M., Clauzon G. & Deaconu F. (2010).- Pliocene and Lower Pleistocene vegetation and climate changes at the European scale: Long pollen records and climatostratigraphy.- Quaternary International, vol. 219, no. 1-2, p. 152-167.
Popescu S.-M., Cavazza W., Suc J.-P., Melinte-Dobrinescu M.C., Barhoun N. & Gorini C. (2021).- Pre-Zanclean end of the Messinian Salinity Crisis: New evidence from Central Mediterranean reference sections.- Journal of the Geological Society, London, vol. 178, no. 3, article jgs2020-183, 16 p.
Popescu S.-M., Suc J.-P. & Loutre M.F. (2006).- Early Pliocene vegetation changes forced by eccentricity-precession. Example from Southwestern Romania.- Palćogeography, Palćoclimatology, Palćoecology, vol. 238, no. 1-4, p. 340-348.
Rehault J.-P., Boillot G. & Mauffret A. (1984).- The western Mediterranean basin geological evolution.- Marine Geology, vol. 55, no. 3-4, p. 447-477.
Robertson A.H.F. & Grasso M. (1995).- Overview of the Late Tertiary-Recent tectonic and palaeoenvironmental development of the Mediterranean region.- Terra Nova, vol. 7, no. 2, p. 114-127.
Rouchy J.-M. (1982).- La genčse des évaporites messiniennes de Méditerrannée.- Mémoires du Muséum national d'Histoire naturelle (Série C, Sciences de la Terre), Paris, vol. 50, 267 p.
Rouchy J.-M. & Caruso A. (2006).- The Messinian salinity crisis in the Mediterranean basin: A reassessment of the data and an integrated scenario.- Sedimentary Geology, vol. 188, p. 35-67.
Rouvier H. (1977).- Géologie de l'extręme nord-tunisien : Tectonique et paléogéographie superposées ŕ l'extrémité orientale de la chaîne nord maghrébine.- Thčse es Sciences, Université Pierre et Marie Curie-Paris VI, 703 p.
Roveri M., Flecker R., Krijgsman W., Lofi J., Lugli S., Manzi V., Sierro F.J., Bertini A., Camer Lenghi A., De Lange G., Govers R., Hilgen F.J., Hübscher C., Meijer P.T. & Stoica M. (2014).- The Messinian Salinity Crisis: Past and future of a great challenge for marine sciences.- Marine Geology, vol. 352, p. 25-58.
Sánchez Gońi M.F., Landais A., Fletcher W.J., Naughton F., Desprat S. & Duprat J. (2008).- Contrasting impacts of Dansgaard-Oeschger events over a western European latitudinal transect modulated by orbital parameters.- Quaternary Science Reviews, vol. 27, no. 11-12, p. 1136-1151.
Saint-Martin J.-P., Hlel S., Debrie J., Ben Moktar N., Saint-Martin S. & Mannaď-Taiech B. (2023).- Microbial influence in the Messinian sedimentation: Example of Cap Bon (NE Tunisia).- Annales de Paléontologie, vol. 109, no. 1, article 102600, 22 p.
Serpelloni E., Vannucci G., Pondrelli S., Argnani A., Casula G., Anzidei M., Baldi B. & Gasperini P. (2007).- Kinematics of the Western Africa-Eurasia plate boundary from focal mechanisms and GPS data.- Geophysical Journal International, vol. 169, no. 3, p. 1180-1200.
Shackleton N.J., Hagelberg T.K. & Crowhurst S.J. (1995a).- Evaluating the success of astronomical tuning: Pitfalls of using coherence as a criterion for assessing pre-Pleistocene timescales.- Paleoceanography, vol. 10, no. 4, p. 693-697.
Shackleton N.J., Hall M.A. & Pate D. (1995b).- Pliocene stable isotope stratigraphy of Site 846. In: Pisias N.G., Mayer L.A., Janecek T.R., Palmer-Julson A. & Andel T.H. van (Eds.), Scientific Results Eastern Equatorial Pacific: Covering Leg 138 of the cruises of the Drilling Vessel JOIDES Resolution, Balboa, Panama, to San Diego, California, Sites 844-854, 1 May-4 July 1991.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 138, p. 337-355.
Spakman W., Chertova M. V., Berg A. van den & Hinsbergen D.J. van (2018).- Puzzling features of western Mediterranean tectonics explained by slab dragging.- Nature Geoscience, vol. 11, no. 3, p. 211-216.
Steininger F.F., Muller C. & Rogl F. (1988).- Correlation of Central Paratethys, Eastern Paratethys, and Mediterranean Neogene stages. In: Royden L.H. & Horvath F. (eds.), The Pannonian Basin, a study in basin evolution.- AAPG Memoir, Boulder - CO, p. 79-87.
Steininger F.F. & Rögl F. (1984).- Paleogeography and palinspastic reconstruction of the Neogene of the Mediterranean and Paratethys.- Geological Society, London, Special Publications, vol. 17, no. 1, p. 659-668.
Suballyova D. (1997, unpublished).- Expression palynologique en Méditerranée des cycles glaciaire-interglaciaire arctiques anciens et cyclostratigraphie : Plio-Pléistocčne de Zakynthos (coupe de la Citadelle) et de Sicile (coupe de Monte San Nicola).- PhD Thesis, University Claude Bernard - Lyon 1, 135 p.
Suc J.-P. (1978).- Analyse pollinique de dépôts plio-pléistocčnes du sud du Massif basaltique de l'Escandorgue (série de Bernasso - Lunas, Hérault - France).- Pollen et Spores, Paris, vol. 20, no. 4, p. 497-512.
Suc J.-P. (1984).- Origin and evolution of the Mediterranean vegetation and climate in Europe.- Nature, vol. 307, no. 5950, p. 429-432.
Suc J.-P. (1989).- Distribution latitudinale et étagement des associations végétales au Cénozoďque supérieur dans l'aire ouest-méditerranéenne.- Bulletin de la Société géologique de France (8e Série), Paris, t. V, no. 3, p. 541-550.
Suc J.-P., Bertini A., Combourieu-Nebout N., Diniz F., Leroy S., Russo-Ermolli E., Zheng Z., Bessais E. & Ferrier J. (1995).- Structure of West Mediterranean vegetation and climate since 5.3 Ma.- Acta zoologica cracoviensia, vol. 38, no. 1, p. 3-16.
Suc J.-P., Clauzon G., Bessedik M., Leroy S., Ambert P. & Martinell J. (1992).- Neogene and lower Pleistocene in Southern France.- Cahiers de Micropaléontologie, Paris, vol. 7, no. 1-2, p. 165-186.
Suc J.-P., Combourieu-Nebout N., Seret G., Popescu S.-M., Klotz S., Gautier F., Clauzon G., Westgate J. & Sandhu A.S. (2010).- The Crotone series: A synthesis and new data.- Quaternary International, vol. 219, no. 1-2, p. 121-133.
Suc J.-P. & Cravatte J. (1982).- Étude palynologique du Pliocčne de Catalogne (nord-est de l'Espagne) : Apports ŕ la connaissance de l'histoire climatique de la Méditerranée occidentale et implications chronostratigraphiques.- Paléobiologie Continentale, Montpellier, vol. XIII, no. 1, p. 1-31.
Suc J.-P. & Drivaliari A. (1991).- Transport of bisaccate coniferous fossil pollen grains to coastal sediments: An example from the earliest Pliocene Orb Ria (Languedoc, Southern France).- Review of Palaeobotany and Palynology, vol. 70, no. 3, p. 247-253.
Suc J.-P., Fauquette S. & Popescu S.-M. (2004).- L'investigation palynologique du Cénozoďque passe par les herbiers. In: Les herbiers : Un outil d'avenir. Tradition et modernité.- Actes du Colloque de Lyon (20-22 novembre 2002), Association Française pour la Conservation des Espčces Végétales, Paris, p. 67-87.
Suc J.-P., Fauquette S., Popescu S.-M. & Robin C. (2020).- Subtropical mangrove and evergreen forest reveal Paleogene terrestrial climate and physiography at the North Pole.- Palćogeography, Palćoclimatology, Palćoecology, vol. 551, article 109755, 9 p.
Suc J.-P., Fauquette S., Warny S., Jiménez-Moreno G. & Do Couto D. (2023).- Climate and Atlantic sea-level recorded in Southwestern Spain from 6.3 to 5.2 Ma. Inferences on the Messinian Crisis in the Mediterranean.- BSGF-Earth Sciences Bulletin, Paris, vol. 194, article 15, 18 p.
Suc J.-P. & Frizon de Lamotte D. (2019).- Paléoenvironnements méditerranéens : Changements et bouleversements environnementaux depuis 23 Ma.- Géochronique, Paris, vol. 151, no. 16-17.
Suc J.-P. & Popescu S.-M. (2005).- Pollen records and climatic cycles in the North Mediterranean region since 2.7 Ma.- Geological Society, London, Special Publications, vol. 247, p. 147-158.
Suc J.-P., Popescu S.-M., Bertini A., Head M.J., Gibbard P.L. & Diniz F. (2018a).- The pioneering contributions and legacy of Waldo H. Zagwijn (1928-2018).- Alpine and Mediterranean Quaternary, Monterotondo, vol. 31, no. 1-2, p. iii-viii.
Suc J.-P., Popescu S.-M., Fauquette S., Bessedik M., Jiménez-Moreno G., Bachiri Taoufiq N., Zheng Z., Medail F. & Klotz S. (2018b).- Reconstruction of Mediterranean flora, vegetation and climate for the last 23 million years based on an extensive pollen dataset.- Ecologia mediterranea, vol. 44, no. 2, p. 53-85.
Suc J.-P. & Zagwijn W.H. (1983).- Plio-Pleistocene correlations between the northwestern Mediterranean region and northwestern Europe according to recent biostratigraphic and palaeoclimatic data.- Boreas, vol. 12, no. 3, p. 153-166.
Svenning J.-C. (2003).- Deterministic Plio-Pleistocene extinctions in the European cool-temperate tree flora.- Ecology Letters, vol. 6, no. 7, p. 646-653.
Targhi S., Barhoun N., Bachiri Taoufiq N., Achab M., Essamoud R., Rahmouna J. & Berry N. (2023).- Late Miocene biostratigraphy and paleoenvironment of the Sais Basin (southern Rifian Corridor, Morocco): New insights from the Moulay Yakoub area.- Carnets Geol., vol. 23, no. 3, p. 53-75. DOI: 10.2110/carnets.2023.2303
Targhi S., Barhoun N., Bachiri Taoufiq N., Achab M., Salem A.A. & Yousfi M.Z. (2021).- Vegetation climate and marine environmental reconstruction in the western Mediterranean (southern Rifian corridor, Morocco) over the Tortonian-Messinian transition.- Heliyon, vol. 7, no. 12, article e08569, 19 p.
Tavarnelli E., Butler R.W.H., Decandia F.A., Calamita F., Grasso M. & Alvarez W. (2004).- Implications of fault reactivation and structural inheritance in the Cenozoic tectonic evolution of Italy. In: Crescenti U., D'Offizi S., Merlini S. & Sacchi R. (eds.), The geology of Italy.- Special Volume of the Italian Geological Society for the IGC 32 Florence-2004, vol. 1, p. 209-222.
Tayech B. (1984).- Études palynologiques dans le Néogčne du Cap-Bon (Tunisie).- Thčse de Doctorat (3e Cycle), Université Claude Bernard-Lyon I, 138 p.
Temani R. & Gaaloul N. (2007).- Les paléopeuplements du Messinien et du Pliocčne du golfe de Hammamet : Intéręt paléoécologique et événementiel.- Revue Méditerranéenne de l'Environnement, Tunis vol. 2, p. 130-142.
Thunell R.C. & Williams D.F. (1983).- The stepwise development of Pliocene-Pleistocene paleoclimatic and paleoceanographic conditions in the Mediterranean: Oxygen isotopic studies of DSDP Sites 125 and 132. In: Meulenkamp J.E. (ed.), Reconstruction of marine paleoenvironments.- Utrecht Micropaleontological Bulletins, vol. 30, p. 111-127.
Torelli L., Grasso M., Mazzoldi G., Peis D. & Gori D. (1995).- Cretaceous to Neogene structural evolution of the Lampedusa shelf (Pelagian Sea, Central Mediterranean).- Terra Nova, vol. 7, no. 2, p. 200-212.
Tzedakis P.C. (2007).- Seven ambiguities in the Mediterranean palaeoenvironmental narrative.- Quaternary Science Reviews, vol. 26, no. 17-18, p. 2042-2066.
Wang C.W. (1961).- The forest of China, with a survey of grassland and desert vegetation.- Maria Moors Cabot Foundation, Publication 5, Harvard University Press, Cambridge - MA, 326 p.
Warny S. (1999, unpublished).- Mio-Pliocene palynology of the Gibraltar Arc: A new perspective on the Messinian Salinity Crisis.- Thčse de Doctorat, Université Catholique de Louvain, 307 p.
Zachariasse W.J., Zijderveld J.D.A., Langereis C.G., Hilgen F.J. & Verhallen P.J.J.M. (1989).- Early late Pliocene biochronology and surface water temperature variations in the Mediterranean.- Marine Micropaleontology, vol. 14, no. 4, p. 339-355.
Zagwijn W.H. (1960).- Aspects of the Pliocene and Early Pleistocene vegetation in the Netherlands.- Mededelingen van de Geologische Stichting (Serie C), Maastricht, III - I - no. 5, 78 p.
Zagwijn W.H. (1998).- Borders and boundaries: A century of stratigraphical research in the Tegelen-Reuver area of Limburg (The Netherlands).- Mededelingen - Nederlands Instituut voor Toegepaste Geowetenschappen TNO, Utrecht, vol. 60, p. 19-34.
Zheng Z. (1990).- Végétations et climats néogčnes des Alpes maritimes franco-italiennes d'aprčs les données de l'analyse palynologique.- Paléobiologie continentale, Montpellier, vol. XVII, p. 217-244.
Zheng Z. & Cravatte J. (1986).- Étude palynologique du Pliocčne de la Côte d'Azur (France) et du littoral ligure (Italie).- Geobios, Villeurbanne, vol. 19, no. 6, p. 815-823.
Plate
1: 1) Taxodium (JR-1, 1140 m, Mega-mesotherm element); 2) Engelhardia (JR-1, 2480 m,
Mega-mesotherm element); 3) Buxus sempervirens type (JR-1, 1400 m, Mesotherm element); 4) Quercus (JR-1, 920 m, Mesotherm element); 5) Cathaya (JR-1, 1140 m); 6-7) Pinaceae (JR-1, 540 m); 8) Cistus (JR-1, 620 m, Mediterranean
xerophyte); 9) Quercus ilex-coccifera type (JR-1, 1840 m, Mediterranean
xerophyte); 10) Olea (JR-1, 1860 m, Mediterranean xerophyte);
(.../...) |
|
Plate
2: 11) Ephedra (JR-1, 680 m, Subdesertic element);
12) Lygeum (JR-1, 640 m, Subdesertic element); 13) Artemisia (JR-1, 640 m, Subdesertic element); 14)
Compositae (Asteraceae) Asteroideae (JR-1, 1820 m, Herb); 15) Compositae
(Asteraceae) Cichorioideae (JR-1, 1760 m, Herb); 16) Brassicaceae (JR-1, 710 m, Herb); 17) Caryophyllaceae (JR-1, 1640 m, Herb); 18) Cyperaceae (JR-1, 1975 m, Herb); 19) Plantago (JR-1, 1240 m, Herb); 20) Poaceae (JR-1, 1300 m, Herb); 21) Amaranthaceae-Chenopodiaceae (JR-1, 660 m, Halophyte). |
|