◄ Carnets Geol. 25 (13) ►
![]()
Outline
[1. Introduction]
[2. Geological and palaeontological framework]
[3. Materials and methods]
[4. Systematic ichnology]
[5.Discussion]
[6.Conclusions] and ...
[Bibliographic references]
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Corso di Dottorato in Geoscienze e Ambiente, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Gruppo Avis Mineralogia e Paleontologia Scandicci, Piazza Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (FI) (Italy);
Istituto di Fisica Applicata “Nello Carrara”, CNR-IFAC, Via Madonna del
Piano 10, 50019 Sesto Fiorentino (FI) (Italy)
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Gruppo Avis Mineralogia e Paleontologia Scandicci, Piazza Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (FI) (Italy)
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Museo di Storia Naturale, Universitŕ di Pisa, via Roma 79, 56011 Calci (PI) (Italy)
Published online in final form (pdf) on November 4, 2025
DOI 10.2110/carnets.2025.2513
![]()
[Editor:
Daniela Basso; language editor: Zev Brook;
technical editor: Bruno R.C. Granier]
![]()
Despite the common and widespread occurrence of turtle shell remains in many fossil vertebrate assemblages worldwide, few palaeontological studies have addressed the palaeoecological significance of turtle bone modification features. Here, we report on ichnofossils occurring on carapacial and plastral bones of the small terrapin genus Mauremys (Testudinoidea, Geoemydidae) from Upper Pliocene deposits exposed at La Serra, in the Valdelsa basin of Tuscany (central Italy). The trace fossil assemblage includes Karethraichnus lakkos, Gunnellichnus cf. moghraensis, and Thatchtelithichnus holmani, all of which are found on the nonvisceral surfaces of the shell. As observed elsewhere, some of the Karethraichnus lakkos borings occur along the epidermal sulci that mark the border between adjacent keratinous scutes. Based on the relevant literature, the studied ichnofossils are interpreted as being due to the parasitic action of leeches/spirorchiid liver flukes (K. lakkos and Thatchtelithichnus holmani) and to algal/fungal/bacterial (sub)cutaneous infections (Gunnellichnus cf. moghraensis). The observed stock of bioerosional features compares favourably with the Italian Pliocene climate, which was warmer and wetter than today.
• palaeoichnology;
• Gunnellichnus moghraensis;
• Karethraichnus lakkos;
• Thatchtelithichnus holmani;
• turtle;
• shell;
• Piacenzian;
• Valdelsa basin
Mulč F., Casati S., Bonfigli L., Di Cencio A. & Collareta A. (2025).- Terrapin aches and pains: Parasite borings and other bone modifications in an Italian Pliocene population of Mauremys (Testudines, Geoemydidae).- Carnets Geol., Madrid, vol. 25, no. 13, p. 265-281. DOI: 10.2110/carnets.2025.2513
Lésions osseuses chez des tortues d'eau douce : Perforations parasitaires et autres modifications osseuses dans une population pliocčne italienne de Mauremys (Testudines, Geoemydidae).- Malgré la présence fréquente et généralisée de restes de carapace de tortue dans de nombreux assemblages fossiles de vertébrés ŕ travers le monde, peu d'études paléontologiques ont abordé la signification paléoécologique des altérations observées sur les os de tortue. Nous rendons compte ici de la découverte d'ichnofossiles observés sur des os de la dossičre et du plastron d'un genre de petite tortue - Mauremys (Testudinoidea, Geoemydidae) - provenant de couches du Pliocčne supérieur affleurant ŕ La Serra, dans le bassin du Val d'Elsa en Toscane (Italie centrale). L'association de traces fossiles comprend Karethraichnus lakkos, Gunnellichnus cf. moghraensis et Thatchtelithichnus holmani, qui se trouvent tous sur les surfaces non viscérales de la carapace. Comme observé ailleurs, certaines des perforations de Karethraichnus lakkos se produisent le long des sillons épidermiques qui marquent la frontičre entre les écailles kératiniques adjacentes. D'aprčs la littérature spécialisée, les ichnofossiles étudiés sont interprétés comme dus ŕ l'action parasitaire de sangsues/douves du foie (K. lakkos et Thatchtelithichnus holmani) et ŕ des infections (sous-)cutanées algales/fongiques/bactériennes (Gunnellichnus cf. moghraensis). L'ensemble des traces bioérosives observées corrobore l'idée que le climat du Pliocčne italien était plus chaud et plus humide qu'aujourd'hui.
• paléoichnologie ;
• Gunnellichnus moghraensis ;
• Karethraichnus lakkos ;
• Thatchtelithichnus
holmani ;
• tortue ;
• carapace ;
• Plaisancien ;
• bassin du Val d'Elsa
Turtles are characterised by possessing a peculiar bony shell that includes both dermal bones and highly modified endoskeletal elements (Rice et al., 2016). This shell comprises of a dorsal, dome-shaped armoured element (i.e., the carapace) and a ventral, flattened one (i.e., the plastron), each of which consists of a mosaic of articulating bony plates. In most cases, the external surfaces of both the plastron and the carapace as well as the periphery of the internal surfaces are coated by a layer of keratinised skin, which in turn is typically divided into nonoverlapping epidermal scutes whose often taxonomically informative patterns are reflected by the occurrence of sulci on the underlying bone. Although these epidermal scutes shelter the turtle shell bone from the surrounding environment, they can be infiltrated and bypassed by a wide array of symbionts, which in turn may result in different bone modifications such as pits, holes and grooves. In addition, algal, fungal and bacterial infections may also damage the shell bones (e.g., M.L. Feldman & M.B. Feldman, 2011; Zonneveld & Bartels, 2019, 2022, 2023; Christiansen et al., 2020; Zonneveld et al., 2022a). The resulting bone modifications appear to be common in modern turtle populations in both aquatic and terrestrial settings and are likely to be frequent in fossil populations as well. That said, little attention has historically been paid by palaeontologists to the ichnology of the turtle shell (with some remarkable exceptions, e.g., Hutchison & Frye, 2001), though this has dramatically changed lately. Among the several works that have dealt with this issue over the last years (e.g., Collareta et al., 2020a, 2022a; Sato & Jenkins, 2020; Collareta & Bosselaers, 2022; Ascarrunz & Joyce, 2024; Guerrero et al., 2024, 2025), those by Zonneveld et al. (2015, 2022b) and Zonneveld and Bartels (2022, 2023) are particularly relevant in the way they have established an updated ichnotaxonomic framework for dealing with such bone modifications and their fossil diversity.
Our aim here is to report on the bone modifications affecting carapacial and plastral bones of Mauremys (Testudinoidea: Geoemydidae) from Upper Pliocene coastal deposits exposed at La Serra (Tuscany, central Italy). The occurrence of Mauremys at this locality was recently assessed based on an incomplete skeleton that also features two examples of the parasite boring Thatchtelithichnus holmani (Collareta et al., 2020a). Including several newly collected plates, the present study reveals a higher diversity of traces, among which are representatives of the ichnogenera Gunnellichnus and Karethraichnus. Some hypotheses on the identity of the tracemakers are proposed, and the palaeobiological and palaeoenvironmental significance of the studied ichnofossils is briefly discussed.
The
fossil specimens discussed herein come from an active quarry in the vicinity of
La Serra village (San Miniato municipality, Pisa Province; geographic
coordinates: 43°39'28"N 10°48'57"E). This quarry
lies in the northwestern sector of the Valdelsa basin, a 25×60 km-wide,
NW-trending extensional basin in the Northern Apennine hinterland (Fig.
1 ![]() ). The
basin fill reaches a maximum thickness of about 2 km, and ranges
chronostratigraphically between the Upper Miocene and the Lower Pleistocene (e.g.,
Abbazzi et al., 2008; Benvenuti
et al., 2014).
). The
basin fill reaches a maximum thickness of about 2 km, and ranges
chronostratigraphically between the Upper Miocene and the Lower Pleistocene (e.g.,
Abbazzi et al., 2008; Benvenuti
et al., 2014).
|
|
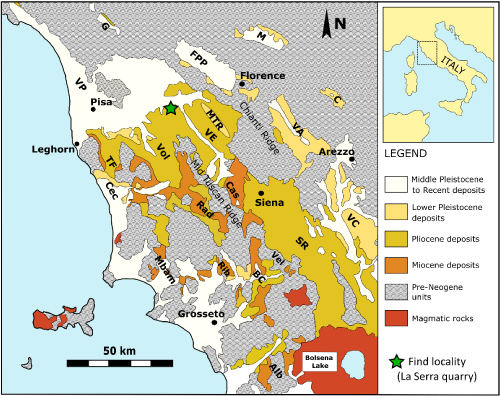
Figure 1: Simplified geological map of
Tuscany (central Italy). The star indicates the geographic position of the La
Serra quarry (GPS geographic coordinates: 43°39'28"N 10°48'57"E) where the terrapin specimens discussed
in the present paper were
discovered. Abbreviations: Alb, Albegna basin; BC, Baccinello–Cinigiano
basin; C, Casentino basin; Cec, Cecina basin; FPP, Firenze–Prato–Pistoia
basin; G, Garfagnana basin; M, Mugello basin; Mbam, Montebamboli basin; MTR,
Montespertoli–Tavernelle ridge; Rad, Radicondoli basin; Rib, Ribolla basin;
SR, Siena–Radicofani basin; TF, Tora–Fine basin; VA, upper Valdarno basin;
VC, Valdichiana basin; VE, Valdelsa–lower Valdarno basin; Vel, Velona basin;
Vol, Volterra basin. Redrawn and modified
from Benvenuti et al. (2014). |
A detailed characterization of the sedimentary succession exposed at La Serra has been provided by Garassino et al. (2012), who recognised the following informal units:
"Unit 1", cropping out in the lower part of the quarry, up to 8 m above the base of the section (hereafter, abs). This unit consists of mixed sandstone and siltstone overlain by cross-stratified sandstone and massive claystone. Garassino et al. (2012) regarded this unit as reflecting an estuarine or deltaic depositional setting punctuated by marshes and ponds;
"Unit 2" (from 8 m to 14 m abs): a thick sandstone succession with high-angle cross-stratifications, interpreted by Garassino et al. (2012) as testifying to a barrier island where both submarine longshore dunes and subaerial aeolian dunes were present;
"Unit 3" (from 14 m to 19.5 m abs): structureless siltstone and silty sandstone that pass laterally into massive claystone. According to Garassino et al. (2012), this unit was deposited in a sheltered, lagoonal environment that was intermittently connected with the sea;
"Unit 4" (from 19.5 m abs to the top of the section, ca. 25 m abs): highly fossiliferous, clayey sandstone that testifies to a low-energy, shallow-marine depositional setting.
Over the last few years, the La Serra quarry has yielded a rich assemblage of terrestrial, freshwater and marine vertebrates. Garassino et al. (2012) were the first to record the occurrence of indeterminate remains of turtles and cervids from Unit 1. Among these finds are plastral and carapacial remains of Mauremys sp., some of which have been described elsewhere (Collareta et al., 2020a). Tortoise remains have also been recently reported from Unit 1, representing the first unambiguous record of the genus Testudo in the Italian Pliocene (Mulč et al., 2025). Unit 2 is home to fragmentary mandibles and long bones of cervids, postcranial bones and a tooth of proboscidean, and cranial and dental elements of suids (Merella et al., 2023). A fairly diverse marine vertebrate fauna comes from Unit 4, including shark teeth as well as the longest stingray stinger ever described from the fossil and modern records (Collareta et al., 2020b). Invertebrate remains occur throughout the succession, with Unit 4 preserving an exceptionally rich decapod fauna (Garassino et al., 2012) as well as the youngest lingulid brachiopods known from the Mediterranean Sea (Di Cencio et al., 2021).
The chronostratigraphic assignment of the aforementioned sediments and fossils is somewhat debated. According to the biostratigraphic results presented by Garassino et al. (2012), the La Serra section should be referred to the Lower Pliocene, with Unit 1 being possibly as old as the uppermost Miocene (Messinian). This interpretation clashes with previous studies that have placed the La Serra section well into the Pliocene (e.g., Benvenuti et al., 1995a, 1995b; Dominici et al., 1995; Abbazzi et al., 2008). Building upon basin-wide correlations, biochronological considerations, and biostratigraphic analyses from many different sites of the Valdelsa basin, this succession was assigned by Dominici et al. (2018) to their S6 Synthem, which in turn would have deposited during the late Piacenzian and early Gelasian. More specifically, Dominici et al. (2018) and Dominici and Danise (2023) assigned Unit 4 of Garassino et al. (2012) to the uppermost Piacenzian, which indicates that the quarry section should be entirely referred to the Upper Pliocene. This chronostratigraphic interpretation appears to be consistent with (i) the Piacenzian age of the underlying S5 Synthem; (ii) the recognition of the Gauss–Matuyama palaeomagnetic transition at Montopoli, c. 6 km from La Serra (Lindsay et al., 1980); and (iii) the subsequent recognition of a simplified nannoplankton assemblage consistent with the CNPL zones 3 (p.p.), 4 and 5 (p.p.), encompassing the upper part of the Zanclean as well as the whole Piacenzian, in the sediments of Unit 4 (Di Cencio et al., 2021). It is also worth noting that the San Miniato area is the source of historical terrestrial vertebrate faunas belonging to the Triversa and Montopoli faunal units, which in turn correspond to the Early–Middle Villafranchian mammal age, embracing the Late Pliocene and earliest Pleistocene (e.g., Rustioni & Mazza, 2001; Rook et al., 2017; Bartolini-Lucenti et al., 2022). In light of the above considerations, we regard the La Serra section as entirely belonging in the Upper Pliocene.
The ichnological nomenclature is mostly derived from Zonneveld et al. (2015, 2022b) and Zonneveld and Bartels (2022, 2023). According to these studies, penetrative traces ("holes") pass through the whole thickness of a given bone, whereas non-penetrative traces ("pits") do not. "Shallow" pits end within the cortical bone layer, whereas "deep" pits penetrate into the internal cancellous bone layer. Individual traces are identified with Greek letters (e.g., "α", "β", etc.).
The fossil specimens discussed herein are housed at Badia a Settimo (Scandicci, Florence Province), in the permanent exhibition of "Gruppo AVIS Mineralogia e Paleontologia Scandicci" (=GAMPS). They consist of one isolated peripheral, one isolated costal, one incomplete plastron and four incomplete epiplastra, originating from a minimum of five turtle individuals.
Specifically, GAMPS-01985 (Fig. 2.e1-2 ![]() ) is a peripheral, whereas GAMPS-01986 is a distal fragment of a
costal (Fig. 2.f1-2
) is a peripheral, whereas GAMPS-01986 is a distal fragment of a
costal (Fig. 2.f1-2 ![]() ). GAMPS-01987 is an isolated right epiplastron, whereas
GAMPS-01967, GAMPS-01983 and GAMPS-01984 are isolated left epiplastra.
GAMPS-01984 and GAMPS-01987 are essentially complete (Fig.
2.a1-2, .b1-2
). GAMPS-01987 is an isolated right epiplastron, whereas
GAMPS-01967, GAMPS-01983 and GAMPS-01984 are isolated left epiplastra.
GAMPS-01984 and GAMPS-01987 are essentially complete (Fig.
2.a1-2, .b1-2 ![]() ),
whereas GAMPS-10967 and GAMPS-10983 lack their posterolateral corners (Fig.
2.c1-2, .d1-2
),
whereas GAMPS-10967 and GAMPS-10983 lack their posterolateral corners (Fig.
2.c1-2, .d1-2 ![]() ). Finally, GAMPS-00925 (see Collareta et al., 2020a: Figs. 2-3) consists of carapacial and plastral bones
from a single turtle individual, including the slightly damaged left and right
hypoplastra as well as the essentially complete left and right xiphiplastra.
). Finally, GAMPS-00925 (see Collareta et al., 2020a: Figs. 2-3) consists of carapacial and plastral bones
from a single turtle individual, including the slightly damaged left and right
hypoplastra as well as the essentially complete left and right xiphiplastra.
|
|
Figure 2: Mauremys sp., newly collected carapacial
and plastral bones featuring bone modifications from the Upper Pliocene of La
Serra. (a1, a2) GAMPS-01987, epiplastron; (b1, b2) GAMPS-01984, epiplastron;
(c1, c2) GAMPS-01983, epiplastron; (d1, d2) GAMPS-01967, epiplastron; (e1, e2)
GAMPS-01985, peripheral, (f1, f2) GAMPS-01986, costal. External views (a1–f1)
and visceral/internal views (a2–f2). All panels are digital renderings of the
textured photogrammetric 3D models. |
All the
aforementioned plates are thin and slender, as usual for freshwater turtles, and
compare favourably with published illustrations and descriptions of the shell
bones of Mauremys (e.g., Chesi, 2009; Chesi et al.,
2009; Colombero et
al., 2017; Georgalis et al., 2019; Vlachos et al.,
2019). In
particular, the epiplastra (Fig. 2.a-d ![]() ) are characterised by a marked anterior
expansion of the area covered by the gular scutes; they display no gular pocket;
and a small, elongated bulge is present medial to the gular-humeral groove (e.g.,
Hervet, 2000). Furthermore, the xiphiplastra (Collareta et
al., 2020a: Fig. 3) exhibit a very deep anal notch and a straight
femoral-anal sulcus.
) are characterised by a marked anterior
expansion of the area covered by the gular scutes; they display no gular pocket;
and a small, elongated bulge is present medial to the gular-humeral groove (e.g.,
Hervet, 2000). Furthermore, the xiphiplastra (Collareta et
al., 2020a: Fig. 3) exhibit a very deep anal notch and a straight
femoral-anal sulcus.
Photographs of all specimens were taken at Dipartimento di Scienze della Terra, Universitŕ di Pisa, using a Nikon D850 camera equipped with a Nikon Micro Nikkor AF-S 60 mm f/2.8 G ED macro-lens and a Zeiss Axio Zoom.V16 macroscope equipped with a Zeiss Axiocam 305 color camera. The latter instrument is connected to a computer running the ZEN core 3.10 software and allows for automated focus stacking as well as for elaborating depth maps and topographic cross-sections over small surfaces.
Textured 3D models of all the trace-bearing bones were elaborated through photogrammetry by using the Agisoft Metashape Standard (64 bit) software. All models were subsequently scaled in Blender 3.6 to their natural size. They are freely available on Figshare at the following web links:
10.6084/m9.figshare.28391534 (GAMPS-00925);
10.6084/m9.figshare.28391642 (GAMPS-01967);
10.6084/m9.figshare.28391615 (GAMPS-01983);
10.6084/m9.figshare.28391651 (GAMPS-01984);
10.6084/m9.figshare.28391663 (GAMPS-01985);
10.6084/m9.figshare.28391666 (GAMPS-01986);
10.6084/m9.figshare.28391675 (GAMPS-01987).
Blender 3.6 was also used to elaborate textured and untextured digital renderings of all models as orthographic views under low-angle light.
Ichnogenus Gunnellichnus Zonneveld et al., 2022b
This ichnogenus consists of shallow, narrow to wide, irregularly etched/corroded areas on the external surface of the shell bones. The outer trace margin is smoothly scalloped to gently undulose, and only rarely straight. Marginal troughs deeper than the centre of the trace do sometimes occur. Gunnellichnus is typically much wider than deep, with a width-to-depth ratio >10:1 (and commonly >100:1). The absolute dimensions of the trace are variable: the examples described by Zonneveld et al. (2022b: Figs. 5–7) vary between 2 mm wide and approximately twice as long to 1–2 cm wide and 3–7 cm long. The included ichnospecies are Gunnellichnus moghraensis (the type ichnospecies) and G. akolouthiste (Zonneveld et al., 2022b).
Gunnellichnus cf. moghraensis Zonneveld et al., 2022b
(Fig. 3 ![]() )
)
Gunnellichnus moghraensis consists of wide surface etchings. The trace depth is typically submillimetric. The outer trace margin is smoothly scalloped to gently undulose. The width-to-depth ratio is >25:1 and commonly exceeds 100:1. The trace floor is often rough and uneven (Zonneveld et al., 2022b).
Fossil occurrences: Gunnellichnus moghraensis was first recognised on plastral bones of the extinct terrapin Erymnochelys from the Lower Miocene of Wadi Moghra, Egypt (Zonneveld et al., 2022b). Collareta and Bosselaers (2022) recorded the presence of Gunnellichnus moghraensis on a cheloniid costal plate from the Middle Miocene of Flanders, Belgium. Janssen et al. (2013) had previously described similar scars on Upper Cretaceous remains of the sea turtle Allopleuron hofmanni from the Maastrichtian type area, ranging across the Netherlands and Belgium. Other occurrences of Gunnellichnus moghraensis are on middle Eocene geoemydid turtles of Wyoming, USA (Zonneveld & Bartels, 2023) as well as on a fragmentary plastron of a 'ptychogasterid' geoemydid from the Eocene of the Messel Pit, in Germany (Ascarrunz & Joyce, 2024).
Material examined herein: "α" and "β", two individual traces on the external surface of epiplastron GAMPS-01987; "γ" and "δ", two individual traces on the internal surface of peripheral GAMPS-01985.
Description:
Two relatively large examples of Gunnellichnus cf. moghraensis
occur on the external surface of GAMPS-01987 (Fig. 3.a1 ![]() ). The partly preserved
trace α stretches along the suture with the adjoining epiplastron (Fig.
3.a2
). The partly preserved
trace α stretches along the suture with the adjoining epiplastron (Fig.
3.a2 ![]() ). Assuming an elliptical outline, the long axis of the trace reconstructs
as parallel to the aforementioned suture. This 5.8-mm-long boring displays
rather regular margins and a relatively smooth floor. Occurring close to the
posterolateral corner of the bone, trace β lies close to the suture with
the adjoining hyoplastron (Fig. 3.a3
). Assuming an elliptical outline, the long axis of the trace reconstructs
as parallel to the aforementioned suture. This 5.8-mm-long boring displays
rather regular margins and a relatively smooth floor. Occurring close to the
posterolateral corner of the bone, trace β lies close to the suture with
the adjoining hyoplastron (Fig. 3.a3 ![]() ). It measures about 5 mm in maximum length
and 3.6 mm in maximum width and penetrates more shallowly than the base of the
cortical bone layer. Trace β displays an elliptical outline having its long
axis parallel to the lateral margin of the bone. This trace is shallow and
provided with a gently scalloped outer margin. A small, irregular, slightly
uplifted boss is observed on the trace floor, its outline recalling that of the
whole pit. Overall, this trace is similar to a G. moghraensis scar
described by Collareta and Bosselaers
(2022: Fig. 4.B, trace "δ"
therein) on a cheloniid costal plate from the Miocene of Belgium.
). It measures about 5 mm in maximum length
and 3.6 mm in maximum width and penetrates more shallowly than the base of the
cortical bone layer. Trace β displays an elliptical outline having its long
axis parallel to the lateral margin of the bone. This trace is shallow and
provided with a gently scalloped outer margin. A small, irregular, slightly
uplifted boss is observed on the trace floor, its outline recalling that of the
whole pit. Overall, this trace is similar to a G. moghraensis scar
described by Collareta and Bosselaers
(2022: Fig. 4.B, trace "δ"
therein) on a cheloniid costal plate from the Miocene of Belgium.
Two larger examples
of Gunnellichnus are present on the internal surface of peripheral
GAMPS-01985, indicated herein as γ
and δ (Fig. 3.b1 ![]() ). The γ
pit lies along the partly damaged suture with an adjoining peripheral (Fig.
3.b2
). The γ
pit lies along the partly damaged suture with an adjoining peripheral (Fig.
3.b2 ![]() ). Measuring about 4.2 mm in length and about 3.7 mm in transverse width, it is wide and shallow, with fairly regular
margins. The trace shape is elliptical, the long axis being parallel to the
aforementioned suture. Near the γ boring, the larger yet incompletely preserved δ pit occurs
close to the outer border of the visceral surface of the bone (Fig.
3.b2
). Measuring about 4.2 mm in length and about 3.7 mm in transverse width, it is wide and shallow, with fairly regular
margins. The trace shape is elliptical, the long axis being parallel to the
aforementioned suture. Near the γ boring, the larger yet incompletely preserved δ pit occurs
close to the outer border of the visceral surface of the bone (Fig.
3.b2 ![]() ). Its
morphology and dimensions are similar to those of trace β on GAMPS-01987.
). Its
morphology and dimensions are similar to those of trace β on GAMPS-01987.
Our Tuscan Pliocene representatives of Gunnellichnus include specimens that are morphologically close to G. moghraensis but typically characterised by a lower width-to-depth ratio than specified in the original diagnosis of Zonneveld et al. (2022b), hence our referral to G. cf. moghraensis. They are nonetheless consistent with similarly sized traces assigned by Zonneveld and Bartels (2023: Fig. 8) to G. moghraensis.
|
|
Figure 3: Gunnellichnus cf. moghraensis borings
on plastral and carapacial bones of Mauremys sp. from the Upper Pliocene
of La Serra. (a1) GAMPS-01987, epiplastron, featuring traces "α" (a2)
and "β" (a3) on its external surface. (b1) GAMPS-01985, peripheral,
featuring traces "γ" and "δ" (b2) on its internal surface.
Panels a1 and b1 are digital renderings of the textured photogrammetric 3D
models. Panels (a2), (a3) and (b2) are photographs taken
under annular light (a2, b2) or low-angle artificial light (a3). |
Ichnogenus Karethraichnus Zonneveld et al., 2015
This ichnogenus is represented by both penetrative holes and non-penetrative pits with a circular or subcircular outline. The latter include shallow, bowl-shaped borings as well as deeper, subcylindrical pits provided with a rounded, blunt, or pointed terminus. The included ichnospecies are Karethraichnus lakkos (the type ichnospecies), K. fiale, K. kulindros, K. minimum, and K. zaratan (Zonneveld et al., 2015; Sato & Jenkins, 2020; Moura et al., 2021). Of all these ichnospecies, only K. fiale conforms to the penetrative morphotype.
Karethraichnus lakkos Zonneveld et al., 2015
This ichnospecies consists of circular, subcircular or irregular pits with a circular outline and a flattened or rounded (i.e., hemispherical) terminus (Zonneveld et al., 2015: Fig. 8.1i, 2022a: Fig. 14.A–C). The traces enter the cortical bone and, in some cases, the shallows of the trabecular bone. The diameter ranges widely between about 1 mm and centimetric, and depth is also variable. These traces are often deeper and more circular on tortoise material than on aquatic turtles (width-to-depth ratio range from 1:1 to 5:1) (Zonneveld et al., 2022b).
Fossil occurrences: Zonneveld et al. (2015) first described this ichnospecies from bony elements of the geoemydid terrapin cf. Echmatemys from the lower Eocene of Wyoming, USA. Collareta et al. (2022a) recorded Karethraichnus cf. lakkos on a cheloniid costal plate from the Upper Miocene of Tuscany (Italy). Zonneveld et al. (2022b) reported on the presence of this ichnospecies on Erymnochelys from the Lower Miocene of Wadi Moghra, Egypt. Collareta et al. (2022a) dealt with two Karethraichnus lakkos borings on a fragmentary sea turtle carapace from the Upper Miocene of the Pisco basin, Peru. Zonneveld and Bartels (2023) recorded this ichnospecies on turtle shell elements of middle Eocene freshwater turtles from Wyoming, USA. El Hedeny et al. (2023) reported on K. lakkos borings on Campanian terrapin materials from Kharga Oasis, Egypt. Szczygielski et al. (2024) detected this ichnospecies on materials of Proterochersis from the Upper Triassic of Baden-Württemberg, Germany. Finally, Ascarrunz and Joyce (2024) recorded Karethraichnus lakkos on a plastral fragment from the Eocene of the Messel Pit, Germany.
Besides turtles, K. lakkos has been recorded on the bones of mosasaurs (as Gastrochaenolites isp., Jagt et al., 2020), megatheriids (Trifilio et al., 2023), and glyptodonts (Zonneveld et al., 2024).
Material examined herein: "ε", "ζ" and "η", three individual traces on the external surface of epiplastron GAMPS-01984; "θ" and "ι", two individual traces on the external surface of epiplastron GAMPS-01983; "κ", "λ" and "μ", three individual traces on the external surface of epiplastron GAMPS-01967; "ν", one individual trace on the external surface of costal GAMPS-01986.
Description:
The small pits ε, ζ and η occur on the external surface of
epiplastron GAMPS-01984, and specifically in its gular region (Fig.
4.a1-3 ![]() ).
Trace ε has a subcylindrical shape and subcircular outline with
well-defined margins (Fig. 4.a2
).
Trace ε has a subcylindrical shape and subcircular outline with
well-defined margins (Fig. 4.a2 ![]() ). The walls of the boring are subvertical, and
its terminus is somewhat lifted above the trace floor (Fig.
4.a3
). The walls of the boring are subvertical, and
its terminus is somewhat lifted above the trace floor (Fig.
4.a3 ![]() ). The diameter
of this trace is somewhat shorter than 2 mm and its depth is about 0.5 mm. Not
far from trace ε, trace ζ is a shallower pit with a hemispherical
shape (Fig. 4.a2
). The diameter
of this trace is somewhat shorter than 2 mm and its depth is about 0.5 mm. Not
far from trace ε, trace ζ is a shallower pit with a hemispherical
shape (Fig. 4.a2 ![]() ). Near the latter lesion, trace η crosses the
gular-humeral sulcus (Fig. 4.a2
). Near the latter lesion, trace η crosses the
gular-humeral sulcus (Fig. 4.a2 ![]() ). It is smaller than the former borings but
similar in shape to trace ε. A few other bone modifications reminiscent of Karethraichnus
lakkos occur on the humeral portion of GAMPS-01984, though these are too
indistinct to be positively identified as trace fossils (Fig.
4.a1
). It is smaller than the former borings but
similar in shape to trace ε. A few other bone modifications reminiscent of Karethraichnus
lakkos occur on the humeral portion of GAMPS-01984, though these are too
indistinct to be positively identified as trace fossils (Fig.
4.a1 ![]() ).
).
|
|
Figure 4: Karethraichnus lakkos borings on
plastral bones of Mauremys sp. from the Upper Pliocene of La Serra. (a1)
GAMPS-01984, epiplastron, featuring traces "ε", "ζ" and "η"
(a2) on its external surface. Panel (a1) is a digital rendering of the textured
photogrammetric 3D model. Panel (a2) is a photograph taken under low-angle
artificial light. Panel (a3) is a cross-section of trace ε, featuring a
somewhat uplifted terminus. |
The small pits θ
and ι occur on the external surface of epiplastron GAMPS-01967 (Fig.
5.a1 ![]() ). The former trace is found close to the inter-epiplastral symphysis. Its
outline is subcircular, with a diameter of approximately 1 mm (Fig.
5.a2
). The former trace is found close to the inter-epiplastral symphysis. Its
outline is subcircular, with a diameter of approximately 1 mm (Fig.
5.a2 ![]() ). Trace
ι has an elliptical outline (Fig. 5.a3
). Trace
ι has an elliptical outline (Fig. 5.a3 ![]() ). It is
a shallow, hemispherical depression with a smooth inner surface.
). It is
a shallow, hemispherical depression with a smooth inner surface.
Three pits are
found on the external surface of epiplastron GAMPS-01967, namely, κ,
λ and μ (Fig. 5.b1-2 ![]() ). Trace κ is deeper than the others,
whereas traces λ and μ are shallow pits occurring close to the gular-humeral
sulcus.
). Trace κ is deeper than the others,
whereas traces λ and μ are shallow pits occurring close to the gular-humeral
sulcus.
|
|
Figure 5: Karethraichnus lakkos borings on plastral bones of Mauremys sp. from
the Upper Pliocene of La Serra. (a1) GAMPS-01983, epiplastron, featuring traces "θ" (a2) and
"ι" (a3) on its external surface. (b1)
GAMPS-01967, epiplastron, featuring traces "κ", "λ" and "μ"
(b2) on its external surface. Panels (a1) and (b1) are digital renderings of the
textured photogrammetric 3D models. Panels (a2), (a3) and (b2) are photographs
taken under annular light (a3) or low-angle artificial light (a2, b2). |
Boring ν
occurs on the external surface of costal GAMPS-01986, along the
vertebral-pleural sulcus (Fig. 6.a1-3 ![]() ). Its diameter is about 1 mm, and its
outline is irregularly subcircular. It is a shallow depression with a flattened
floor.
). Its diameter is about 1 mm, and its
outline is irregularly subcircular. It is a shallow depression with a flattened
floor.
All the studied specimens of Karethraichnus lakkos conform to the "Type I" borings of Zonneveld et al. (2022a: Fig. 14.A), i.e., "shallow (<1 mm) circular pits etched into the external cortex". Trace ε may be an exception, as its 3D morphology is similar to the "Type III" of Zonneveld et al. (2022a: Fig. 14.C), i.e., "flat-bottomed pits that terminate within the trabecular layer", although it does not penetrate the trabecular bone due to its limited depth.
|
|
Figure 6: Karethraichnus lakkos borings on
carapacial bones of Mauremys sp. from the Upper Pliocene of La Serra.
(a1) GAMPS-01986, costal, featuring trace "v" (a2, 3) on its external
surface. Panel (a1) is a digital rendering of the textured photogrammetric 3D
model. Panels (a2) and (a3) are photographs taken under annular light (a2) or
low-angle artificial light (a3). |
Ichnogenus Thatchtelithichnus Zonneveld et al., 2015
This monotypic ichnogenus is represented by circular to subcircular, ring-shaped grooves that affect the external layer of the cortical bone. The signature character of these borings lies in the presence of an uplifted central element on the trace floor, which consists of an unmodified pedestal or a rounded nubbin. The trace walls are rounded, convex or sharp, and perpendicular to the substrate (Zonneveld et al., 2015: Fig. 8.2iv-v, 2022a: Fig. 14.F). The trace may (or may not) penetrate the outer part of the trabecular bone. Its diameter is variable, ranging between 1.5–2 mm on Eocene terrapins from Wyoming, USA (Zonneveld et al., 2015) and 7 mm on Miocene freshwater turtles from Wadi Moghra, Egypt (Zonneveld et al., 2022b).
Thatchtelithichnus holmani Zonneveld et al., 2015
(Fig. 7 ![]() )
)
Fossil occurrences: Zonneveld et al. (2015) first described this ichnospecies from the external surface of plastral bones of the geoemydid turtles cf. Echmatemys from the lower Eocene of Wyoming (USA). Zonneveld and Bartels (2019) reassigned to Thatchtelithichnus holmani some rosette-like traces occurring on dinosaur long bones from the Jurassic of Wyoming. Two records reported on by Collareta et al. (2020a) are rediscussed herein. Zonneveld et al. (2022b) referred to T. holmani some pits observed on the plastron bottom of the terrapin Erymnochelys from the Lower Miocene of Wadi Moghra to Thatchtelithichnus.
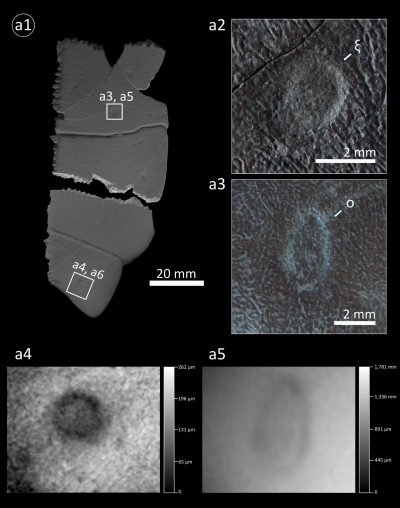
Material examined herein: "ξ" and "o," two individual traces on the external (i.e. ventral) surface of partial plastron GAMPS-00925.
Description: Collareta et
al. (2020a) described two shallow, ring-shaped scars occurring on the
external surface of the partial plastron GAMPS-00925. These pits are refigured
herein with the support of macroscope imaging (Fig. 7.a1-5 ![]() ). Trace ξ,
a boring observed in the abdominal area of the left hypoplastron (Fig.
7.a2, .a4
). Trace ξ,
a boring observed in the abdominal area of the left hypoplastron (Fig.
7.a2, .a4 ![]() ),
is "a very shallow, circular depression, measuring ~3 mm in diameter, hosting
at its centre a raised, rounded nubbin having a diameter of ~1.5 mm" (Collareta
et al., 2020a). The second scar, trace
o, lies in the anal portion of the left
xiphiplastron (Fig. 7.a3, .a5
),
is "a very shallow, circular depression, measuring ~3 mm in diameter, hosting
at its centre a raised, rounded nubbin having a diameter of ~1.5 mm" (Collareta
et al., 2020a). The second scar, trace
o, lies in the anal portion of the left
xiphiplastron (Fig. 7.a3, .a5 ![]() ). It is
"a rather elongate depression, measuring
~4×2.5 mm, with a central unmodified platform that extends for a distance of
less than 1 mm from the outer edge of the trace" (Collareta et
al., 2020a).
). It is
"a rather elongate depression, measuring
~4×2.5 mm, with a central unmodified platform that extends for a distance of
less than 1 mm from the outer edge of the trace" (Collareta et
al., 2020a).
|
|
Figure 7: Thatchtelithichnus holmani borings on
plastral bones of Mauremys sp. from the Pliocene of La Serra. (a1)
GAMPS-00925, left hypoplastron and xiphiplastron, featuring traces "ξ"
(a2) and "o" (a3) on their external surface. Panels a1 is a digital
rendering of the textured photogrammetric 3D model. Panels (a2) and (a3) are
photographs taken under low-angle artificial light. Panels (a4) and (a5) are
depth maps of traces ξ and o, respectively. |
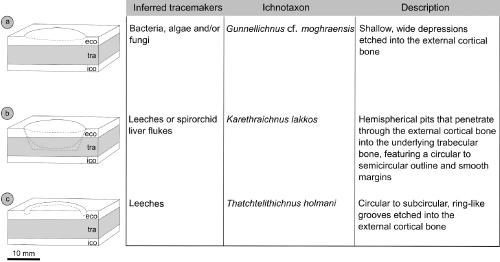
Although the aetiology of the pits and holes observed on modern and fossil carapacial and plastral bones remains somewhat conjectural, it has been noted elsewhere that turtles are subjected to a broad spectrum of ecto- and mesoparasites, among which the producers of these bone modifications should be searched for (e.g., M.L. Feldman & M.B. Feldman, 2011; Christiansen et al., 2020; Zonneveld & Bartels, 2022, 2023). That all the traces dealt with in the present paper occur on the nonvisceral surfaces of the bony elements strongly suggests that trace production occurred due to some external agent while the host turtles were alive (Zonneveld et al., 2015, 2022b; Zonneveld & Bartels, 2022, 2023: table 2).
The three ichnotaxa
recognised on the Tuscan Pliocene shell bones of Mauremys appear to be
relatively widespread among extinct and extant turtles, although the studies
dealing specifically with these issues remain few in number. Gunnellichnus
moghraensis occurs both on Mesozoic and Cenozoic materials as well as
both on sea turtle and terrapin shell bones. Janssen
et al. (2013) proposed that these
bone modifications may be the result of anchoring by a variety of sessile,
epibiotic macro-invertebrates. However, this interpretation has been argued
against by several recent studies (Zonneveld et al.,
2015,
2022b; Collareta & Bosselaers,
2022;
Zonneveld & Bartels, 2022,
2023; Guerrero
et al., 2024), which rather
proposed that Gunnellichnus-like traces are due to algal, bacterial and/or
fungal infections of the turtle shell. Enlightening comparisons have been
recently established with the lesions observed on modern freshwater turtles such
as the red-eared terrapin, Trachemys scripta elegans (Zonneveld
& Bartels, 2022,
2023). Several diseases affecting primarily the
epidermal scutes – e.g., carapacial shell disease (CSD), necrotizing
scute disease (NSD), pond turtle shell disease (PTSD), ulcerative shell disease
(USD) and septicemic cutaneous ulcerative disease (SCUD) – can expand to
eventually damage the shell bones in the form of more or less amorphous
degradation patches (e.g., Lovich et
al., 1996; Garner et al.,
1997; Granados et al., 2013; Rothschild
et al., 2013; Christiansen et
al., 2020). These lesions may be obliterated by subsequent pathological
reaction features (sensu Bertling et
al., 2006, 2022) (Fig. 8.a ![]() ).
).
|
|
Figure 8:
Synoptic overview of the ichnological diversity observed on carapacial and
plastral bones of Mauremys from the Upper Pliocene of La Serra. The
descriptions and sketches of Karethraichnus lakkos and Thatchtelithichnus
holmani are modified after Zonneveld et al.
(2022b: Fig. 14.a). Abbreviations: eco, external cortex;
tra,
trabecular bone; ico, internal cortex. |
As for Karethraichnus lakkos and Thatchtelithichnus holmani, different groups of ecto- and mesoparasites have been proposed as the most likely tracemakers, depending on the hosts' lifestyle and environmental preferences. These include ixodid arachnids (ticks) as well as epizoic barnacles, which are thought to be prominent producers of the aforementioned traces on terrestrial and marine turtles, respectively (Collareta et al., 2022b; Zonneveld et al., 2022a). In freshwater habitats, leeches are the most commonly reported parasites of terrapins (Zonneveld & Bartels, 2022). When feeding, leeches attach themselves to their host with the posterior sucker, whereas a proboscis is extended from the oral opening of the anterior sucker and inserted into the turtle shell (e.g., Siddal & Gaffney, 2004; Fediras et al., 2017). Feeding is accompanied by the release of salivary secretions that may be capable of bone decalcification and digestion of the connective tissue matrix (Siddal & Gaffney, 2004), hence the leeches' ichnogenetic potential (Zonneveld & Bartels, 2022). Similarly, some vermiform parasites such as the spirorchid liver flukes may also cause lesions on the carapace and plastron of aquatic turtles (e.g., Johnson et al., 1998).
The feeding sites of extant leeches are often located in-between adjacent keratinous scutes (e.g., Siddal & Gaffney, 2004; Fediras et al., 2017). This is consistent with the observation of Zonneveld et al. (2015, 2022b) that Karethraichnus lakkos traces are especially frequent along the epidermal sulci. We were able to replicate this observation on the Tuscan Pliocene epiplastron GAMPS-01984 (trace η) as well as on the costal GAMPS-01986 (trace ν).
Regardless of their taxonomic assignment, the main tracemakers of K.
lakkos and Thatchtelithichnus holmani on turtle bones were
likely parasites that prefer plastrons to carapaces, because these borings are
more commonly observed on plastral materials. Plastrons provide a more sheltered
location, which suggests that the tracemakers may be sensitive to exposure or
desiccation (Zonneveld & Bartles,
2022, 2023)
(Fig. 8.b-c ![]() ).
).
According to the palaeoenvironmental reconstruction proposed by Garassino et al. (2012), the Pliocene population of Mauremys discussed herein would have inhabited a coastal lagoon system not far from a river mouth and sheltered seaward by a chain of barrier islands. This is consistent with the broad habitat preferences of modern Mauremys spp., which include brackish water bodies such as coastal lagoons (e.g., Sidis & Gasith, 1985; Ernst & Barbour, 1989; Highfield, 2002; Vamberger et al., 2014).
In the present-day Mediterranean region, Mauremys is represented by M. leprosa, which inhabits the coasts of Northwestern Africa as well as in the Iberian Peninsula (Keller & Busak, 2001), and by M. rivulata, which inhabits the Eastern Mediterranean (Barth et al., 2004). No native populations of Mauremys exist to date on the Apennine Peninsula, from which this genus was extirpated during the Pleistocene (Chesi, 2009; Chesi et al., 2009). Previously, in Pliocene times, Mauremys inhabited Italy under a warmer and more humid climate than today (e.g., Albianelli et al., 1997; Bertini, 2010; Combourieu-Nebout et al., 2015; Prista et al., 2015). These palaeoenvironmental conditions may correlate with the stock of bioerosional features observed on the Mauremys shell remains from La Serra. In particular, diseases affecting the epidermal scutes have been proposed to be sensitive to changes in temperature, rainfall frequency and cloud cover as well as to vitamin deficiency (Zonneveld & Bartels, 2023). Nowadays, Gunnellichnus-like shell lesions are especially frequent on freshwater turtle populations inhabiting persistently moist settings, where fungal and bacterial infections do not dry out easily (e.g., Rose et al., 2001; Paré & Jacobson, 2007) because of a low basking efficacy (Zonneveld & Bartels, 2023).
We reported on the occurrence of different bone modifications on newly collected plastral and carapacial bones of the small terrapin, Mauremys sp., from Upper Pliocene deposits exposed at La Serra, a recently discovered palaeontological site in Pisa Province (Tuscany, central Italy). The recognised ichnotaxa include representatives of Gunnellichnus cf. moghraensis, Karetharichnus lakkos, and Thatchtelithichnus holmani. All the studied traces occur on the nonvisceral surfaces of the bones. As previously observed elsewhere, some Karetharichnus lakkos borings occur along the epidermal sulci that mark the border between adjacent keratinous scutes. Based on the relevant literature, the studied ichnofossils are interpreted as due to algal, bacterial and/or fungal infections (Gunnellichnus cf. moghraensis) as well as to the parasitic action of leeches and/or spirorchid liver flukes (Karetharichnus lakkos and Thatchtelithichnus holmani). A similar suite of traces has recently been reported from the shell bones of the extant red-eared slider, Trachemys scripta elegans, evoking similar tracemakers as well as a comparable ecology for our Tuscan Pliocene population of Mauremys. Previous studies have reconstructed the Mauremys-bearing strata at La Serra as comprising a brackish lagoon system, which matches well with the broad spectrum of environmental preferences of extant Mauremys spp. Our Tuscan Pliocene population of Mauremys lived under warm, humid conditions, which compares favourably with the observed stock of bioerosional features.
Our gratitude goes to Giovanni Bianucci, Marco Merella, Alice Pieri, and Giulia Bosio for sharing several days of fieldwork at La Serra. Thanks are also due to Tassos Kotsakis for fruitful discussions on the systematic affinities of the turtle shell remains from this newly discovered fossil site. Not least, we are sincerely indebted to the journal editors, Daniela Basso, Bruno Granier, and Zev Brook, as well as to Massimo Delfino and an anonymous colleague, whose thorough reviews greatly contributed to improve an early draft of this paper.
Abbazzi L., Benvenuti M., Ceci M.E., Esu D., Faranda C., Rook L. & Tangocci F. (2008).- The end of the Lago-Mare time in the SE Valdelsa Basin (Central Italy): Interference between local tectonism and regional sea-level rise.- Geodiversitas, Paris, vol. 30, p. 611-639.
Albianelli A., Azzaroli A., Bertini A., Ficcarelli G., Napoleone G. & Torre D. (1997).- Paleomagnetic and palynologic investigations in the upper Valdarno basin (central Italy): calibration of an early Villafranchian fauna.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 103, p. 111-118.
Ascarrunz E. & Joyce W.G. (2024).- A plastron fragment reveals a previously unrecorded turtle species in the Eocene of Messel Pit, Germany.- Fossil Record, Berlin, vol. 27, p. 259-264.
Barth D., Bernhard D., Fritzsch G. & Fritz U. (2004).- The freshwater turtle genus Mauremys (Testudines, Geoemydidae) - a textbook example of an east-west disjunction or a taxonomic misconcept?. - Zoologica Scripta, London, vol. 33, p. 213-221.
Bartolini-Lucenti S., Cirilli O., Pandolfi L. & Rook L. (2022).- The Montopoli site, a reference local fauna in the Pliocene and Pleistocene European large Mammals biochronology, first discovered by Giovanni Capellini (1833–1922).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 61, p. 207-222.
Benvenuti M., Bertini A., Conti C., Dominici S. & Falcone D. (1995b).- Analisi stratigrafica e paleoambientale integrata del Pliocene dei dintorni di San Miniato.- Quaderni del Museo di Storia Naturale di Livorno, Livorno, vol. 14, p. 29-49.
Benvenuti M., Del Conte S., Scarselli N. & Dominici S. (2014).- Hinterland basin development and infilling through tectonic and eustatic processes: Latest Messinian-Gelasian Valdelsa Basin, Northern Apennines, Italy.- Basin Research, vol. 26, p. 387-402.
Benvenuti M., Dominici S. & Rook L. (1995a).- Inquadramento stratigrafico-deposizionale delle faune a mammiferi villafranchiane (Unitŕ Faunistiche Triversa e Montopoli) del Valdarno inferiore nella zona a sud dell'Arno (Toscana).- Il Quaternario, Roma, vol. 8, p. 457-464.
Bertini A. (2010).- Pliocene to Pleistocene palynoflora and vegetation in Italy: State of the art.- Quaternary International, vol. 225, p. 5-24.
Bertling M., Braddy S.J., Bromley R.G., Demathieu G.R., Genise J., Mikuláš R., Nielsen J.K., Nielsen K.S.S., Rindsberg A.K., Schlirf M. & Uchman A. (2006).- Names for trace fossils: A uniform approach.- Lethaia, Stockholm, vol. 39, p. 265-286.
Bertling M., Buatois L. A., Knaust D., Laing B., Mángano M.G., Meyer N., Mikuláš R., Minter N.J., Neumnann C., Rindsberg A. K., Uchman A. & Wisshak M. (2022).- Names for trace fossils 2.0: Theory and practice in ichnotaxonomy.- Lethaia, Stockholm, 55, p. 1-19.
Chesi F. (2009).- Il registro fossile italiano dei cheloni.- Ph.D. Thesis, Universitŕ degli Studi di Firenze, Florence, 178 p.
Chesi F., Delfino M. & Rook L. (2009).- Late Miocene Mauremys (Testudines, Geoemydidae) from Tuscany (Italy): Evidence of terrapin persistence after a mammal turnover.- Journal of Paleontology, Lawrence - KS, vol. 83, p. 379-388.
Christiansen J.L., Davis D.R., Jacobsen E.R. & Laduc T.J. (2020).- Carapacial shell disease process revealed by a long-term field study of the mud turtle, Kinosternon flavescens, in Texas.- Journal of Herpetology, Columbus - OH, vol. 54, p. 1-8.
Collareta A. & Bosselaers M. (2022).- A new record of Gunnellichnus moghraensis from the Middle Miocene of Belgium, with some remarks on the origin of this seemingly uncommon ichnospecies.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 305, p. 237-243.
Collareta A., Casati S., Zuffi M.A.L. & Di Cencio A. (2020a).- First authentic record of the freshwater turtle Mauremys from the Upper Pliocene of Italy, with a new occurrence of the rarely reported ichnotaxon Thatchtelithichnus holmani.- Carnets Geol., Madrid, vol. 20, no. 16, p. 301-313. DOI: 10.2110/carnets.2020.2016
Collareta A., Merella M., Bosselaers M., Casati S., Di Cencio A. & Bianucci G. (2022a).- A Karethraichnus boring on a turtle shell bone from the Miocene of Italy is assessed as the attachment scar of a platylepadid symbiont.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 303, p. 327-337.
Collareta A., Merella M., Casati S. & Di Cencio A. (2020b).- Did titanic stingrays wander the Pliocene Mediterranean Sea? Some notes on a giant-sized myliobatoid stinger from the Piacenzian of Italy.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 298, p. 155-164.
Collareta A., Varas-Malca R., Bosio G., Urbina M. & Coletti G. (2022b).- Ghosts of the Holobiont: Borings on a Miocene turtle carapace from the Pisco Formation (Peru) as witnesses of ancient symbiosis.- Journal of Marine Science and Engineering, vol. 11, no. 1, article 45, 16 p.
Colombero S., Alba D.M., D'Amico C., Delfino M., Esu D., Giuntelli P., Harzhauser M., Mazza P.P.A., Mosca M., Neubauer T.A., Pavia G., Pavia M., Villa A. & Carnevale G. (2017).- Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications.- Palaeontologia Electronica, Los Angeles - CA, vol. 20, article 20.1.10A, p. 1-66. DOI: 10.26879/658
Combourieu-Nebout N., Bertini A., Russo-Ermolli E., Peyron O., Klotz S., Montade V., Fauquette S., Allen J., Fusco F., Goring S., Huntlay B., Joannin S., Lebreton V., Magri D., Martinetto E., Orain R. & Sadori L. (2015).- Climate changes in the central Mediterranean and Italian vegetation dynamics since the Pliocene.- Review of Palaeobotany and Palynology, vol. 218, p. 127-147.
Di Cencio A., Dulai A., Catanzariti R., Casati S. & Collareta A. (2021).- First record of the brachiopod Lingula? From the Pliocene of Tuscany (Italy): The youngest occurrence of lingulides in the Mediterranean Basin.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 299, p. 237-249.
Dominici S. & Danise S. (2023).- Mediterranean onshore–offshore gradient in the composition and temporal turnover of benthic molluscs across the middle Piacenzian Warm Period.- Geological Society, London, Special Publications, vol. 529, p. 365-394.
Dominici S., Danise S. & Benvenuti M. (2018).- Pliocene stratigraphic paleobiology in Tuscany and the fossil record of marine megafauna.- Earth-Science Reviews, vol. 176, p. 277-310.
Dominici S., Mazzanti R. & Nencini C. (1995).- Geologia dei dintorni di San Miniato tra l'Arno, l'Elsa e l'Era.- Quaderni del Museo di Storia Naturale di Livorno, vol. 14, p. 1-35.
El Hedeny M., Mohesn S., Tantawy A.A., El-Sabbagh A., AbdelGawad M. & El-Kheir G.A. (2023).- Bioerosion traces on the Campanian turtle remains: New data from the lagoonal deposits of the Quseir Formation, Kharga Oasis, Egypt.- Palaeontologia Electronica, Los Angeles - CA, vol. 26, article 26.3.a40, p. 1-21. DOI: 10.26879/1315
Ernst C.H., Barbour R.W. & Altenburg R.G.M. (1989).- Turtles of the World.- Smithsonian Institution Press, Washington - DC, p. 1432-1521.
Fediras S., Rouag R., Ziane N., Olivier A., Béchet A. & Benyacoub S. (2017).- Prevalence of Placobdella costata (Fr. Műller, 1846) (Hirudinida: Glossiphoniidae) on the European pond turtle (Emys orbicularis) in northeast Algeria.- Herpetology Notes, Columbus - OH, vol. 10, p. 3-8.
Feldman M.L. & Feldman M.B. (2011).- A tale of two shell diseases: Observations of two shell diseases present in breeding operations of the red-eared slider turtle (Trachemys scripta elegans).- Turtle and Tortoise Newsletter, New York – NY, vol. 15, p. 25-29.
Garassino A., Pasini G., De Angeli A., Charbonnier S., Famiani F., Baldanza A. & Bizzarri R. (2012).- The decapod community from the Early Pliocene (Zanclean) of La Serra quarry (San Miniato, Pisa, Toscana, central Italy): Sedimentology, systematics, and palaeoenvironmental implications.- Annales de Paléontologie, Paris, vol. 98, p. 1-61.
Garner M.M., Herrington R., Howerth E.W., Homer B.L., Nettles V.F., Izara R., Shotts E.B. Jr. & Jacobsen E.R. (1997).- Shell disease in river cooters (Pseudemys concinna) and yellow-bellied turtles (Trachemys scripta) in a Georgia (USA) lake.- Journal of Wildlife Diseases, Leesburg - VA, vol. 33, p. 78-86.
Georgalis G., Villa A., Ivanov M., Vasilyan D. & Delfino M. (2019).- Fossil amphibians and reptiles from the Neogene locality of Maramena (Greece), the most diverse European herpetofauna at the Miocene/Pliocene transition boundary.- Palaeontologia Electronica, Los Angeles - CA, vol. 22, article 22.3.68, p. 1-99. DOI: 10.26879/908
Granados J.L, Moreno O.G. & Brieva C.I. (2013).- Lesiones ulcerativas cutáneas en tortugas dulceacuícolas.- Revista de la Facultad de Medicina Veterinaria y de Zootecnia, Bogotá, vol. 60, p. 61-70.
Guerrero A., Ortega F., Martín de Jesús S. & Pérez-García A. (2024).- Postcranial anomalies of Eocene freshwater pleurodiran and cryptodiran turtles from the Spanish Duero Basin.- The Anatomical Record, Glendale, p. 1-18.
Guerrero A., Smith T. & Pérez-García A. (2025).- Bioerosional marks in the shells of two sea turtle taxa from the middle Eocene of Belgium.- Fossil Record, Berlin, vol. 28, p. 45-56.
Hervet S. (2000).- Tortues du Quaternaire de France : Critčres de détermination, répartitions chronologique et géographique.- Mésogée, Marseille, vol. 58, p. 3-47.
Highfield A.C. (2002).- An introduction to the Mauremys turtles of the Mediterranean.- Tortoise Trust online article, available from: http://www.tortoisetrust.org/articles/maurem ys.html
Hutchison J.H. & Frye F.L. (2001).- Evidence of pathology in early Cenozoic turtles.- PaleoBios, Berkeley - CA, vol. 21, p. 12-19.
Jagt J.W., Deckers M.J., De Leebeeck M., Donovan S.K. & Nieuwenhuis E. (2020).- Episkeletozoans and bioerosional ichnotaxa on isolated bones of Late Cretaceous mosasaurs and cheloniid turtles from the Maastricht area, the Netherlands.- Geologos, Poznań, vol. 26, p. 39-49.
Janssen R., Baal R.R. van & Schulp A.S. (2013).- Bone damage in Allopleuron hofmanni (Cheloniidae, Late Cretaceous).- Netherlands Journal of Geosciences, Utrecht, vol. 92, p. 153-157.
Johnson C.A., Griffith J.W., Tenorio P., Hytrek S. & Lang C.M. (1998).- Fatal trematodiasis in research turtles.- Comparative Medicine, Memphis, vol. 48, p. 340-343.
Keller C. & Busack S.D. (2001).- Mauremys leprosa (Schweigger, 1812) - Maurische Bachschildkröte.- Handbuch der Reptilien und Amphibien Europas, Strasbourg, vol. 3, p. 57-88.
Lindsay E.H., Opdyke N.O. & Johnson N.M. (1980).- Pliocene dispersal of the horse Equus and late Cenozoic mammalian dispersal events.- Nature, vol. 287, p. 135-138.
Lovich J.E., Gotte S.W., Ernst C.H., Harshbarger J.C., Laemmerzahl A.F. & Gibbons J.W. (1996).- Prevalence and histopathology of shell disease in turtles from Lake Blackshear, Georgia.- Journal of Wildlife Diseases, Leesburg, vol. 32, p. 259-265.
Merella M., Collareta A., Casati S., Di Cencio A. & Bianucci G. (2023).- Pliocene geotourism: Innovative projects for valorizing the paleontological heritage of three different-staged quarries of Tuscany (Central Italy).- Geoheritage, vol. 15, article 82, 17 p.
Moura J.F., Nascimento C.S.I., Peixoto B. de C.P. e M., Barros G.E.B. de, Robbi B. & Fernandes M.A. (2021).- Damaged armour: Ichnotaxonomy and paleoparasitology of bioerosion lesions in osteoderms of Quaternary extinct armadillos.- Journal of South American Earth Sciences, vol. 109, Article 103255, 17 p.
Mulč F., Casati S., Bonfigli L., Di Cencio A. & Collareta A. (2025).- First unambiguous record of the tortoise genus Testudo from the Italian Pliocene.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 316, article 1287, 15 p.
Paré J.A. & Jacobson E.R. (2007).- Mycotic diseases of reptiles. In: Jacobson E.R. (eds.), Infectious Diseases and Pathology of Reptiles.- CRC Press, Boca Raton - FL, p. 527-570.
Prista G.A., Agostinho R.J. & Cachăo M.A. (2015).- Observing the past to better understand the future: A synthesis of the Neogene climate in Europe and its perspectives on present climate change.- Open Geosciences, Berlin, vol. 7, p. 65-83.
Rice R., Kallonen A., Cebra-Thomas J. & Gilbert S.F. (2016).- Development of the turtle plastron, the order-defining skeletal structure.- Proceedings of the National Academy of Sciences, Washington - DC, vol. 113, p. 5317-5322.
Rook L., Cirilli O. & Bernor R.L. (2017).- Una forma tardiva di "Hipparion" nell'associazione faunistica del Villafranchiano medio di Montopoli, Italia (Pleistocene inferiore; MN16b; ca. 2.5 Ma).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 56, p. 333-339.
Rose F.L., Koke J., Koehn R. & Smith D. (2001).- Identification of the etiological agent for necrotizing scute disease in the Texas Tortoise.- Journal of Wildlife Diseases, Leesburg, vol. 37, p. 223-228.
Rothschild B.M., Schultze H.-P. & Pellegrini R. (2013).- Chapter 27. Osseous and other tissue pathologies in turtles and abnormalities in mineral deposition. In: Brinkman D.B., Holroyd P.A. & Gardner J.D. (eds.), Morphology and evolution of turtles.- Springer, p. 501-534.
Rustioni M. & Mazza P. (2001).- Taphonomic analysis of Tapirus arvernensis remains from the lower Valdarno (Tuscany, Central Italy).- Geobios, Villeurbanne, vol. 34, p. 469-474.
Sato K. & Jenkins R.G. (2020).- Mobile home for pholadoid boring bivalves: First example from a Late Cretaceous sea turtle in Hokkaido Japan.- Palaios, Lawrence - KS, vol. 35, p. 228-236.
Siddall M.E. & Gaffney E.S. (2004).- Observations on the leech Placobdella ornata feeding from bony tissues of turtles.- Journal of Parasitology, Lawrence - KS, vol. 90, p. 1186-1188.
Sidis I. & Gasith A. (1985).- Food habits of the Caspian terrapin (Mauremys caspica rivulata) in unpolluted and polluted habitats in Israel.- Journal of Herpetology, Columbus - OH, vol. 19, p. 108-115.
Szczygielski T., Marchetti L. & Dróżdż D. (2024).- Natural external plastron mold of the Triassic turtle Proterochersis: An unusual mode of preservation.- PLoS ONE, vol. 19, article e0299314, 30 p. DOI: 10.1371/journal.pone.0299314
Trifilio L.H.M. da S., Araújo Júnior H.I. de & Porpino K. de O. (2023).- The paleoichnofauna in bones of Brazilian quaternary cave deposits and the proposition of two new ichnotaxa.- Ichnos, Philadelphia - PA, vol. 30, p. 207-234.
Vamberger M., Stuckas H., Ayaz D., Lymberakis P., Široký P. & Fritz U. (2014).- Massive transoceanic gene flow in a freshwater turtle (Testudines: Geoemydidae: Mauremys rivulata).- Zoologica Scripta, vol. 43, p. 313-322.
Vlachos E., Sterli J., Vasileiadou K. & Syrides G. (2019).- A new species of Mauremys (Testudines, Geoemydidae) from the late Miocene–Pliocene of Central Macedonia (northern Greece) with exceptionally wide vertebral scutes.- Papers in Palaeontology, vol. 5, p. 177-195.
Zonneveld J.-P., AbdelGawad M.K. & Miller E.R. (2022b).- Ectoparasite borings, mesoparasite borings, and scavenging traces in early Miocene turtle and tortoise shell: Moghra Formation, Wadi Moghra, Egypt.- Journal of Paleontology, Lawrence - KS, vol. 96, p. 304-322.
Zonneveld J.-P. & Bartels W.S. (2019).- Ichnologic note: In defence of Thatchtelithichnus Zonneveld, Bartels, Gunnell and McHugh, 2015.- Ichnos, Philadelphia - PA, vol. 27, p. 152-255.
Zonneveld J.-P. & Bartels W.S. (2022).- The occurrence of bone modification features in the carapace and plastron of the extant red-eared slider Trachemys scripta elegans (Wied-Neuwied, 1839): Implications for paleoecological analyses of fossil turtle assemblages.- Palaios, Lawrence - KS, vol. 37, p. 499-519.
Zonneveld J.-P. & Bartels W.S. (2023).- The occurrence and significance of Ulcerative Shell Disease (USD) on geoemydid turtles, Middle Eocene, Southwestern Wyoming, United States.- Palaios, Lawrence - KS, vol. 38, p. 453-473.
Zonneveld J.-P., Bartels W.S., Gunnell G.F. & McHugh L.P. (2015).- Borings in early Eocene turtle shell from the Wasatch Formation, South Pass, Wyoming.- Journal of Paleontology, Lawrence - KS, vol. 89, p. 802-820.
Zonneveld J.-P., Wilson O. & Holroyd P. (2024).- Composite ichnological-pathological evidence for arthropod parasitism on osteoderms of Boreostemma acostae (Glyptodontidae, Cingulata) from La Venta, Colombia.- Journal of South American Earth Sciences, vol. 146, article 105085, 13 p.
Zonneveld J.-P., Zonneveld Z.E.E., Bartels W.S., Gingras M.K. & Head J.J. (2022a).- Bone modification features resulting from barnacle attachment on the bones of loggerhead sea turtles (Caretta caretta), Cumberland Island, Georgia, USA: Implications for the paleoecological, and taphonomic analyses of fossil sea turtles.- Palaios, Lawrence - KS, vol. 37, p. 650-670.